También podría gustarte
- 0141 (1) EeDocumento4 páginas0141 (1) EeMariana MansillaAún no hay calificaciones
- Accretion Profile of The Rosy Barb, Puntius Conchonius (Hamilton-Buchanan, 1822) From The Water Bodies of Nagaland, IndiaDocumento3 páginasAccretion Profile of The Rosy Barb, Puntius Conchonius (Hamilton-Buchanan, 1822) From The Water Bodies of Nagaland, IndiaInternational Organization of Scientific Research (IOSR)Aún no hay calificaciones
- A Description of The Growth of Sheep and Its Genetic AnalysisDocumento13 páginasA Description of The Growth of Sheep and Its Genetic AnalysisYessenia FigueroaAún no hay calificaciones
- G2910 Heritability and Its Use in Animal Breeding - University of Missouri ExtensionDocumento8 páginasG2910 Heritability and Its Use in Animal Breeding - University of Missouri ExtensionAlbyziaAún no hay calificaciones
- Evaluation of Alternative Schemes For Recording Body Weights in Meat-Type Quails by Using Random RegressionDocumento7 páginasEvaluation of Alternative Schemes For Recording Body Weights in Meat-Type Quails by Using Random RegressionLuciano Pinheiro da SilvaAún no hay calificaciones
- Length-Weight Relationship and Condition Factor of Channa Aurantimaculata (Musikasinthorn, 2000) Studied in A Riparian Wetland of Dhemaji District, Assam, IndiaDocumento6 páginasLength-Weight Relationship and Condition Factor of Channa Aurantimaculata (Musikasinthorn, 2000) Studied in A Riparian Wetland of Dhemaji District, Assam, IndiaresearchinbiologyAún no hay calificaciones
- Biometric Models in Animal BreedingDocumento12 páginasBiometric Models in Animal BreedingRamachandran Ram100% (1)
- Artikel TwinDocumento6 páginasArtikel TwinMuch MahmudiAún no hay calificaciones
- Using Factor Scores in Multiple Linear Regression Model For Predicting The Carcass Weight of Broiler Chickens Using Body MeasurementsDocumento5 páginasUsing Factor Scores in Multiple Linear Regression Model For Predicting The Carcass Weight of Broiler Chickens Using Body MeasurementsVedika AryaAún no hay calificaciones
- Genetic Evaluation of Crossbred Cattle For Reproduction TraitsDocumento8 páginasGenetic Evaluation of Crossbred Cattle For Reproduction TraitsTJPRC PublicationsAún no hay calificaciones
- Bize Oecologia2002Documento4 páginasBize Oecologia2002richnerAún no hay calificaciones
- 5 Kecermatan Dugaan Respon Selektif Bobot Badan PDFDocumento8 páginas5 Kecermatan Dugaan Respon Selektif Bobot Badan PDFramdanAún no hay calificaciones
- The Relationship Between Body Condition Score and Reproductive PerformanceDocumento8 páginasThe Relationship Between Body Condition Score and Reproductive PerformanceL MarsiniAún no hay calificaciones
- Estimation of Genetic and Phenotypic Parameters For Sow Productivity Traits in South African Large White PigsDocumento9 páginasEstimation of Genetic and Phenotypic Parameters For Sow Productivity Traits in South African Large White Pigsonurb90Aún no hay calificaciones
- 1 s2.0 S0022030203738946 MainDocumento10 páginas1 s2.0 S0022030203738946 MainAlondraAún no hay calificaciones
- MC KayDocumento4 páginasMC Kayanamia28Aún no hay calificaciones
- Nature 02104Documento4 páginasNature 02104Ricardo Esquivel SilvaAún no hay calificaciones
- 1 s2.0 S0032579119321170 MainDocumento7 páginas1 s2.0 S0032579119321170 MainGuilherme Costa VenturiniAún no hay calificaciones
- Cow Weight in A Closed Brahman HerdDocumento4 páginasCow Weight in A Closed Brahman HerdKattie ValerioAún no hay calificaciones
- PHD ArticleDocumento5 páginasPHD ArticlebhakthaAún no hay calificaciones
- ProjectreportDocumento31 páginasProjectreportapi-515961562Aún no hay calificaciones
- Abstarct الخلاصة انجليزيDocumento3 páginasAbstarct الخلاصة انجليزيjasim ali sammerAún no hay calificaciones
- (68-75) Study On Fecundity and GSI of Carassius Carassius Linneaus Introduced) From Dal Lake KashmirDocumento9 páginas(68-75) Study On Fecundity and GSI of Carassius Carassius Linneaus Introduced) From Dal Lake KashmirAlexander DeckerAún no hay calificaciones
- 1 s2.0 S0737080611001559 Main PDFDocumento2 páginas1 s2.0 S0737080611001559 Main PDFLaviniaMenicucciAún no hay calificaciones
- Animal - BreedingDocumento34 páginasAnimal - Breedingvignesh murugesanAún no hay calificaciones
- Author's Accepted Manuscript: Livestock ScienceDocumento15 páginasAuthor's Accepted Manuscript: Livestock ScienceDomínguez Derik EriksenkAún no hay calificaciones
- Birth Weight and Biometry of Purebred Landrace Pigs Under Indian Farm ConditionDocumento5 páginasBirth Weight and Biometry of Purebred Landrace Pigs Under Indian Farm Conditionskmdec09Aún no hay calificaciones
- Fan R. - Linkage and Association Studies of QTL For Nuclear Families by Mixed Models (2003)Documento21 páginasFan R. - Linkage and Association Studies of QTL For Nuclear Families by Mixed Models (2003)hfjsdhfAún no hay calificaciones
- Fek. Panjang PDFDocumento9 páginasFek. Panjang PDFyusufnabhanAún no hay calificaciones
- Productivity of Taggar Goats As Affected by Sex of Kids and Litter SizeDocumento6 páginasProductivity of Taggar Goats As Affected by Sex of Kids and Litter SizeKeep CalmAún no hay calificaciones
- Biometrika Trust, Oxford University Press BiometrikaDocumento15 páginasBiometrika Trust, Oxford University Press BiometrikanempaAún no hay calificaciones
- Andersen E.W. - Composite Likelihood and Two-Stage Estimation in Family Studies (2004)Documento16 páginasAndersen E.W. - Composite Likelihood and Two-Stage Estimation in Family Studies (2004)matin mehradAún no hay calificaciones
- J Anim Sci (2007) .Volumen85:984-996 Voluntary IntakeDocumento1 páginaJ Anim Sci (2007) .Volumen85:984-996 Voluntary IntakeDavid Alejandro Camargo BaracaldoAún no hay calificaciones
- Genetics and Breeding Genetic Evaluation of Dairy Cattle Using Test Day Yields and Random Regression ModelDocumento10 páginasGenetics and Breeding Genetic Evaluation of Dairy Cattle Using Test Day Yields and Random Regression ModeldomAún no hay calificaciones
- Fetal Scapular LengthDocumento5 páginasFetal Scapular LengthApexAún no hay calificaciones
- 1 s2.0 S0304401702001942 MainDocumento16 páginas1 s2.0 S0304401702001942 Mainyujiao MaAún no hay calificaciones
- The Effect of Body Size On Animal AbundanceDocumento8 páginasThe Effect of Body Size On Animal AbundanceChan ChibiAún no hay calificaciones
- Umbilical Artery Pulsatility Index and Fetal AbdominalDocumento5 páginasUmbilical Artery Pulsatility Index and Fetal AbdominalerasmohuertasAún no hay calificaciones
- Genetic Parameters: P G E ADocumento8 páginasGenetic Parameters: P G E AInnovation IndiaAún no hay calificaciones
- Chapter 6 - Correlation AnalysisDocumento132 páginasChapter 6 - Correlation AnalysisNicole AgustinAún no hay calificaciones
- Log Linear and Logistic Modeling of DepeDocumento18 páginasLog Linear and Logistic Modeling of DepeIsma IAún no hay calificaciones
- tmpE9EC TMPDocumento9 páginastmpE9EC TMPFrontiersAún no hay calificaciones
- 7a. Concept of HeritabilityDocumento12 páginas7a. Concept of HeritabilityNishant ShahAún no hay calificaciones
- Factors Related To The Etiology of Retained Placenta in Dairy CattleDocumento12 páginasFactors Related To The Etiology of Retained Placenta in Dairy CattleNovericko Ginger BudionoAún no hay calificaciones
- Ecology, Physiology, Allometry and Dimensional@: TrevorDocumento6 páginasEcology, Physiology, Allometry and Dimensional@: TrevorMspamAún no hay calificaciones
- Confidence Intervals For Ratio of Two Poisson Rates Using The Method of Variance Estimates RecoveryDocumento23 páginasConfidence Intervals For Ratio of Two Poisson Rates Using The Method of Variance Estimates RecoveryJaneAún no hay calificaciones
- Acta Obstet Gynecol Scand - 2005 - JohnsenDocumento9 páginasActa Obstet Gynecol Scand - 2005 - Johnsenahmed haddadAún no hay calificaciones
- Jurnal Kambing PEDocumento3 páginasJurnal Kambing PEMohammad Ridwan SetiyonoAún no hay calificaciones
- Strong Evidence For An Intraspecific Metabolic Scaling Coefficient Near 0.89 in FishDocumento17 páginasStrong Evidence For An Intraspecific Metabolic Scaling Coefficient Near 0.89 in FishTiberiu MarianAún no hay calificaciones
- Assessmentof Fetal Lungmaturity:Lecithin/ Sphingomyelinratiovs. Cortisoland Foam-Test AssaysDocumento4 páginasAssessmentof Fetal Lungmaturity:Lecithin/ Sphingomyelinratiovs. Cortisoland Foam-Test AssaysPatrick RamosAún no hay calificaciones
- 10 3168@jds 2009-2720 en IdDocumento14 páginas10 3168@jds 2009-2720 en IdUswatun HasanahAún no hay calificaciones
- MetilDocumento7 páginasMetilAmalia Ashd Al-islamAún no hay calificaciones
- Length Weight Relationship and Condition Factor of Puntius Sophore (Hamilton) From Kolkata and Suburban Fish MarketDocumento5 páginasLength Weight Relationship and Condition Factor of Puntius Sophore (Hamilton) From Kolkata and Suburban Fish MarketDR. BIJAY KALI MAHAPATRAAún no hay calificaciones
- Conception Dan Calving Interval Sapi Potong Peranakan OngoleDocumento6 páginasConception Dan Calving Interval Sapi Potong Peranakan OngolefrankyAún no hay calificaciones
- 8816 19063 1 SMDocumento6 páginas8816 19063 1 SMSamuelAún no hay calificaciones
- Climate ChangeDocumento5 páginasClimate ChangeAhsan MunirAún no hay calificaciones
- Fetal PII002822Documento11 páginasFetal PII002822Ophay Rusty'sAún no hay calificaciones
- The Waltham Book of Human-Animal Interaction: Benefits and Responsibilities of Pet OwnershipDe EverandThe Waltham Book of Human-Animal Interaction: Benefits and Responsibilities of Pet OwnershipI. RobinsonAún no hay calificaciones
- Nutrition Conference for Feed Manufacturers: University of Nottingham, Volume 7De EverandNutrition Conference for Feed Manufacturers: University of Nottingham, Volume 7Aún no hay calificaciones
- BHHVKDocumento13 páginasBHHVKTristan ArellanoAún no hay calificaciones
- Appresoria PDFDocumento3 páginasAppresoria PDFrachnnaaAún no hay calificaciones
- Behaviour and Ecology of Spiders: Carmen Viera Marcelo O. Gonzaga EditorsDocumento448 páginasBehaviour and Ecology of Spiders: Carmen Viera Marcelo O. Gonzaga EditorsMojano De la MagdalenaAún no hay calificaciones
- Chamelion With An AttitudeDocumento11 páginasChamelion With An AttitudeRaj KumarAún no hay calificaciones
- APEX The Organ SystemsDocumento171 páginasAPEX The Organ Systemsjt100% (2)
- Yoga FellatioDocumento145 páginasYoga Fellatiorian4me80% (30)
- Literasi Bhs Ing Part 4Documento4 páginasLiterasi Bhs Ing Part 4Fajar Rahman AprialdiAún no hay calificaciones
- Rajkumar Biology Printable Notes Unit 2 by Rajat.21-37Documento17 páginasRajkumar Biology Printable Notes Unit 2 by Rajat.21-37Michael Benton100% (1)
- Austin Is BattyDocumento3 páginasAustin Is BattyAbdulrahmanAún no hay calificaciones
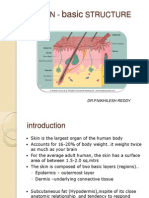
- Skin Structure and DevelopmentDocumento45 páginasSkin Structure and DevelopmentNikhileshReddyAún no hay calificaciones
- Rajkumar Biology Unit - 5 by RajatDocumento52 páginasRajkumar Biology Unit - 5 by RajatShrikant KumarAún no hay calificaciones
- Evolution of Dermal Skeleton and Dentition in Vertebrates The Odontode Regulation TheoryDocumento82 páginasEvolution of Dermal Skeleton and Dentition in Vertebrates The Odontode Regulation TheoryspotxAún no hay calificaciones
- EF3e Preint Filetest 09a Answer SheetDocumento2 páginasEF3e Preint Filetest 09a Answer SheetTanweer ElnaggarAún no hay calificaciones
- Katayama&Endo - 2010 - Redescription of A Sanddiver, T. Blochii With Confirmation of Its ValidityDocumento10 páginasKatayama&Endo - 2010 - Redescription of A Sanddiver, T. Blochii With Confirmation of Its ValidityMuntasir AkashAún no hay calificaciones
- WFC 111 Lecture Syllabus 2013Documento2 páginasWFC 111 Lecture Syllabus 2013mrhower_262391642Aún no hay calificaciones
- Biological (Met)Documento52 páginasBiological (Met)SirLhitz B. UmaliAún no hay calificaciones
- 1-5 1 Cell TransportDocumento3 páginas1-5 1 Cell Transportapi-262368188Aún no hay calificaciones
- Caring For Orphan Wild Baby Birds - Rescuing Them, Raising Them, Their Food & CareDocumento21 páginasCaring For Orphan Wild Baby Birds - Rescuing Them, Raising Them, Their Food & CareFaye HowardAún no hay calificaciones
- EvolutionDocumento26 páginasEvolutionapi-262368188Aún no hay calificaciones
- An Introduction To Wolf SpidersDocumento4 páginasAn Introduction To Wolf SpidersJohn GamesbyAún no hay calificaciones
- 8.introduction To EmbryologyDocumento63 páginas8.introduction To EmbryologyAhmed OrabyAún no hay calificaciones
- Unit - V Chapter-22. Excretory Products and Their EliminationDocumento15 páginasUnit - V Chapter-22. Excretory Products and Their EliminationBattuJayakrishnaAún no hay calificaciones
- BIG X Paleontology PDFDocumento434 páginasBIG X Paleontology PDFWach Ijocho Naggx Babetare'Aún no hay calificaciones
- Quagga: From Wikipedia, The Free EncyclopediaDocumento10 páginasQuagga: From Wikipedia, The Free Encyclopediaaquel1983Aún no hay calificaciones
- Maynard Smith - BibliographyDocumento7 páginasMaynard Smith - BibliographyMarkvN2Aún no hay calificaciones
- CLS Aipmt 18 19 XIII Zoo Study Package 3 SET 1 Chapter 12Documento30 páginasCLS Aipmt 18 19 XIII Zoo Study Package 3 SET 1 Chapter 12Saravanan Raju100% (1)
- Marshmalien LabDocumento9 páginasMarshmalien LabShonnefarrow14Aún no hay calificaciones
- June 2015 Question Paper 31Documento20 páginasJune 2015 Question Paper 31LAKSHMANANKISHOREAún no hay calificaciones
- Paper 20 Damyanti Patel PDFDocumento6 páginasPaper 20 Damyanti Patel PDFPRABHAT KUMAR CHAUHANAún no hay calificaciones
- Of Headless Mice... and Men: by Charles KrauthammerDocumento2 páginasOf Headless Mice... and Men: by Charles KrauthammerEvan QuenlinAún no hay calificaciones