También podría gustarte
- Coral Reefs: Ecosystems of Environmental and Human ValueDocumento8 páginasCoral Reefs: Ecosystems of Environmental and Human ValueNurul Huda 귀여운Aún no hay calificaciones
- Causes of Biodiversity Loss in Coastal EcosystemsDocumento12 páginasCauses of Biodiversity Loss in Coastal EcosystemsSHEAN GAYLE ANGAún no hay calificaciones
- Threats To The Coastal ZoneDocumento9 páginasThreats To The Coastal ZoneJames bondAún no hay calificaciones
- Belize Barrier Reef Reserve System - 2020 COA - enDocumento13 páginasBelize Barrier Reef Reserve System - 2020 COA - enbluwashere2Aún no hay calificaciones
- Final PPT MeDocumento18 páginasFinal PPT Meswats387Aún no hay calificaciones
- Marine Pollution Bulletin: Ana Carolina Grillo, Thayn A Jeremias MelloDocumento7 páginasMarine Pollution Bulletin: Ana Carolina Grillo, Thayn A Jeremias MelloSanghita BiswasAún no hay calificaciones
- The Effects of Climate Change On Coral Reefs Evaluation EssayDocumento6 páginasThe Effects of Climate Change On Coral Reefs Evaluation EssayJohnny DinhAún no hay calificaciones
- FormalfinalDocumento8 páginasFormalfinalapi-620232040Aún no hay calificaciones
- Case Study PDFDocumento7 páginasCase Study PDFnarayananAún no hay calificaciones
- Ecosystem-Based Adaptation in Marine Ecosystems - Hale Et Al 2009 RRJ Vol25#4Documento8 páginasEcosystem-Based Adaptation in Marine Ecosystems - Hale Et Al 2009 RRJ Vol25#4cdrewsAún no hay calificaciones
- GBR Best Practice Literature ReviewDocumento14 páginasGBR Best Practice Literature ReviewAliciaAún no hay calificaciones
- Understanding Coastal ProcessesDocumento31 páginasUnderstanding Coastal ProcessesRirin lestariAún no hay calificaciones
- 23, IVAR DO SUL Plastic Pollution at A Sea Turtle Conservation Area in NE Brazil - Contrasting Developed and Undeveloped BeachesDocumento10 páginas23, IVAR DO SUL Plastic Pollution at A Sea Turtle Conservation Area in NE Brazil - Contrasting Developed and Undeveloped BeachesAnonymous HijNGQtNAún no hay calificaciones
- Marine Pollution Bulletin: Brigitte Gavio, Shelly Palmer-Cantillo, J. Ernesto ManceraDocumento13 páginasMarine Pollution Bulletin: Brigitte Gavio, Shelly Palmer-Cantillo, J. Ernesto ManceraPaola CárdenasAún no hay calificaciones
- Interference With Coastal Structures: o o o oDocumento6 páginasInterference With Coastal Structures: o o o oROSALIE AGMALIWAún no hay calificaciones
- Interference With Coastal Structures: o o o oDocumento6 páginasInterference With Coastal Structures: o o o oROSALIE AGMALIWAún no hay calificaciones
- R R R Roooocccck K K Ky y y y Sssshhhhoooorrrreeee H H H Ha Aa Abbbbiiiittttaaaatttt Aaaaccccttttiiiioooon N N N Ppppllllaaaan N N NDocumento5 páginasR R R Roooocccck K K Ky y y y Sssshhhhoooorrrreeee H H H Ha Aa Abbbbiiiittttaaaatttt Aaaaccccttttiiiioooon N N N Ppppllllaaaan N N NalidaeiAún no hay calificaciones
- Assessment of The Effects of Keta Sea Defence Project On The Livelihoods of The People in The Keta Municipality, Volta Region, GhanaDocumento13 páginasAssessment of The Effects of Keta Sea Defence Project On The Livelihoods of The People in The Keta Municipality, Volta Region, GhanaPremier PublishersAún no hay calificaciones
- Article 77 - Coastline ErosionDocumento5 páginasArticle 77 - Coastline ErosionAniruddha SoniAún no hay calificaciones
- Biodiversity Index of Corals in Bitaug StA. Cruz Davao Del SurDocumento74 páginasBiodiversity Index of Corals in Bitaug StA. Cruz Davao Del SurMicky BanderaAún no hay calificaciones
- Grim Revelations On The World Coral Reefs EcosystemsDocumento3 páginasGrim Revelations On The World Coral Reefs EcosystemsAneesh AnandAún no hay calificaciones
- Coastal Ecosystems and ResourcesDocumento22 páginasCoastal Ecosystems and Resourcesasha cornellAún no hay calificaciones
- Protection From Coastal ErosionDocumento30 páginasProtection From Coastal ErosionArunrajAún no hay calificaciones
- TSDC Biorock Aff-2Documento40 páginasTSDC Biorock Aff-2Jose CruzAún no hay calificaciones
- How To Fix Broken ChairDocumento1 páginaHow To Fix Broken ChairGuinerAún no hay calificaciones
- White PagesDocumento13 páginasWhite Pagesapi-581906330Aún no hay calificaciones
- A Review of Some Environmental Issues Affecting Marine MiningDocumento14 páginasA Review of Some Environmental Issues Affecting Marine Mininggigio marinoAún no hay calificaciones
- Reef Health and The Aquarium Industry How Aquariums Benefit ReefsDocumento3 páginasReef Health and The Aquarium Industry How Aquariums Benefit ReefsTINALEETNT723Aún no hay calificaciones
- Coral Reef Destruction from Careless TourismDocumento14 páginasCoral Reef Destruction from Careless TourismKimberly CalimlimAún no hay calificaciones
- Introduction 2Documento12 páginasIntroduction 2Jean Denis TavoueAún no hay calificaciones
- MCCIP 2013ARC Backingpapers 18 CHab IIIDocumento13 páginasMCCIP 2013ARC Backingpapers 18 CHab IIIROHIT AGRAWALAún no hay calificaciones
- Biodisel 25Documento22 páginasBiodisel 25thevaraja karuppiaAún no hay calificaciones
- The Importance of Coral Reefs ResearchDocumento8 páginasThe Importance of Coral Reefs ResearchemmAún no hay calificaciones
- Blue Carbon PDFDocumento24 páginasBlue Carbon PDFmohammadyunus1992Aún no hay calificaciones
- Islands White Paper - Copy 2Documento7 páginasIslands White Paper - Copy 2api-711033918Aún no hay calificaciones
- Mangrosana; Future World; Rising Tides, Sinking Islands & the Role of Mangrove Trees: Neurosana, #4De EverandMangrosana; Future World; Rising Tides, Sinking Islands & the Role of Mangrove Trees: Neurosana, #4Aún no hay calificaciones
- Ocean & Coastal ManagementDocumento17 páginasOcean & Coastal ManagementbagepAún no hay calificaciones
- Importance of Coral ReefsDocumento15 páginasImportance of Coral ReefsKallie FokkerAún no hay calificaciones
- VA Report For The Municipality of San JoseDocumento35 páginasVA Report For The Municipality of San JoseRolly Balagon CaballeroAún no hay calificaciones
- Climate Change and Marine Turtles: ReviewDocumento18 páginasClimate Change and Marine Turtles: ReviewThe Science Exchange International Sea Turtle InternshipsAún no hay calificaciones
- Tenses of English in 2014Documento11 páginasTenses of English in 2014Aye XaAún no hay calificaciones
- Reef Degradation and Coral Loalversll Y in Indonesia. Effects of Land-Based Pollution, Destructive Fishing Practices and Changes Over TimeDocumento14 páginasReef Degradation and Coral Loalversll Y in Indonesia. Effects of Land-Based Pollution, Destructive Fishing Practices and Changes Over TimeAndiHaerulAún no hay calificaciones
- Marine Sustainability Threatened by Overfishing, Tourism, PollutionDocumento11 páginasMarine Sustainability Threatened by Overfishing, Tourism, PollutionSharlene PenningtonAún no hay calificaciones
- Protection From Coastal ErosionDocumento30 páginasProtection From Coastal ErosionHaryo ArmonoAún no hay calificaciones
- Marine PollutionDocumento22 páginasMarine PollutionAwal ZainalAún no hay calificaciones
- Coral Remote SensingDocumento15 páginasCoral Remote SensingRiskahMappileAún no hay calificaciones
- 13-MENDEZ-Condición Del Arrecife Coralino de Playa BlancaDocumento14 páginas13-MENDEZ-Condición Del Arrecife Coralino de Playa BlancammendezbioAún no hay calificaciones
- Synthesis: Thematic Paper: Synthesis of The Protective Functions of Coastal Forests and Trees Against Natural HazardsDocumento10 páginasSynthesis: Thematic Paper: Synthesis of The Protective Functions of Coastal Forests and Trees Against Natural HazardsHaryo ArmonoAún no hay calificaciones
- Beach Erosion Research PaperDocumento6 páginasBeach Erosion Research Papergzrvpcvnd100% (1)
- Impacts of Drilling On The Outer Continental ShelfDocumento2 páginasImpacts of Drilling On The Outer Continental ShelfMarcus Skookumchuck VanniniAún no hay calificaciones
- Largest Analysis of Coral Reef Health Ever UndertakenDocumento5 páginasLargest Analysis of Coral Reef Health Ever UndertakenagungmalayantapirAún no hay calificaciones
- Review On Coral BleachingDocumento7 páginasReview On Coral Bleachingelissa.pelsmaekersAún no hay calificaciones
- Vietnam's Experience Protecting Coral Reefs for Sustainable DevelopmentDocumento5 páginasVietnam's Experience Protecting Coral Reefs for Sustainable DevelopmentAzra khanAún no hay calificaciones
- Varit's Capstone Project PaperDocumento19 páginasVarit's Capstone Project PaperVarit SuriyasomboonAún no hay calificaciones
- Ocean and Coastal Management: Nelson Rangel-Buitrago, Victor N. de Jonge, William NealDocumento10 páginasOcean and Coastal Management: Nelson Rangel-Buitrago, Victor N. de Jonge, William NealBaluAún no hay calificaciones
- 2.0 Coastal Erosion: Georges Disaster Mitigation (PGDM) Project. The Coastal Erosion Hazard Mapping Presented inDocumento14 páginas2.0 Coastal Erosion: Georges Disaster Mitigation (PGDM) Project. The Coastal Erosion Hazard Mapping Presented inAndi Azizah FebiyantiAún no hay calificaciones
- Coastal erosion mapping aids development planningDocumento14 páginasCoastal erosion mapping aids development planningAndi Azizah FebiyantiAún no hay calificaciones
- Seagrasses Status and Trends in NRM CountriesDocumento34 páginasSeagrasses Status and Trends in NRM CountriesLiHaruAún no hay calificaciones
- Adlerian Psychotherapy: Prioritizing RelationshipsDocumento24 páginasAdlerian Psychotherapy: Prioritizing RelationshipsJayanth MamundiAún no hay calificaciones
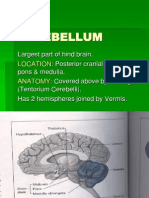
- Physiology of Cerebellum Lec Foe 2nd Year by DR Sadia Uploaded by ZaighamDocumento61 páginasPhysiology of Cerebellum Lec Foe 2nd Year by DR Sadia Uploaded by ZaighamZaigham HammadAún no hay calificaciones
- 皮膚病介紹Documento93 páginas皮膚病介紹MK CameraAún no hay calificaciones
- Biomimicry Avjeet SinghDocumento21 páginasBiomimicry Avjeet SinghAvjeet SinghAún no hay calificaciones
- Muscle Energy Techniques ExplainedDocumento2 páginasMuscle Energy Techniques ExplainedAtanu Datta100% (1)
- 02 Anatomy and Histology PLE 2019 RatioDocumento69 páginas02 Anatomy and Histology PLE 2019 RatioPatricia VillegasAún no hay calificaciones
- (PDF) Root Growth of Phalsa (Grewia Asiatica L.) As Affected by Type of CuttDocumento5 páginas(PDF) Root Growth of Phalsa (Grewia Asiatica L.) As Affected by Type of CuttAliAún no hay calificaciones
- The "Five Families" College Essay ExampleDocumento1 páginaThe "Five Families" College Essay ExampleKishor RaiAún no hay calificaciones
- LWT - Food Science and TechnologyDocumento8 páginasLWT - Food Science and TechnologyAzb 711Aún no hay calificaciones
- Cell Membrane and Transport ColoringDocumento3 páginasCell Membrane and Transport ColoringTeresa GonzAún no hay calificaciones
- Jurnal Perkecambahan Biji PDFDocumento14 páginasJurnal Perkecambahan Biji PDFKinad Danik50% (2)
- Crop Sci 1 Lecture Manual OverviewDocumento86 páginasCrop Sci 1 Lecture Manual OverviewAnalYn Litawan Bucasan100% (1)
- Module 2 NCM 114Documento11 páginasModule 2 NCM 114Erven AranasAún no hay calificaciones
- Summary of Qualitative Tests (Pharmacognosy)Documento8 páginasSummary of Qualitative Tests (Pharmacognosy)kidsaintfineAún no hay calificaciones
- HHS Public Access: Structure and Function of The Human Skin MicrobiomeDocumento20 páginasHHS Public Access: Structure and Function of The Human Skin MicrobiomeEugeniaAún no hay calificaciones
- Microbiological Test of Medical DevicesDocumento5 páginasMicrobiological Test of Medical Devicesbijendra_sinhaAún no hay calificaciones
- Protochordata FIXDocumento33 páginasProtochordata FIXSylvia AnggraeniAún no hay calificaciones
- When The Koel SingsDocumento15 páginasWhen The Koel SingsAlfie Paduga BeguinaAún no hay calificaciones
- Food Chain QsDocumento3 páginasFood Chain QsOliver SansumAún no hay calificaciones
- Natamycin Story - What You Need to KnowDocumento13 páginasNatamycin Story - What You Need to KnowCharles MardiniAún no hay calificaciones
- 1.4 - Diffusion and Osmosis - CrosswordPuzzleDocumento1 página1.4 - Diffusion and Osmosis - CrosswordPuzzlemmssthAún no hay calificaciones
- JCM 08 00217 v3Documento23 páginasJCM 08 00217 v3Sumit BediAún no hay calificaciones
- Pemphigus Subtypes Clinical Features Diagnosis andDocumento23 páginasPemphigus Subtypes Clinical Features Diagnosis andAnonymous bdFllrgorzAún no hay calificaciones
- Biomedical Physics, Vol 7 @medphyslibDocumento360 páginasBiomedical Physics, Vol 7 @medphyslibStats -MANAún no hay calificaciones
- Physiological Psychology ReviewerDocumento13 páginasPhysiological Psychology ReviewerLeopando Rod CyrenzAún no hay calificaciones
- Artigo Sobre Nutrigenômica e Nutrigenética PDFDocumento18 páginasArtigo Sobre Nutrigenômica e Nutrigenética PDFDiogo JuniorAún no hay calificaciones
- HistamineDocumento16 páginasHistaminevg03ik2Aún no hay calificaciones
- Amazing Facts - Exercises 1Documento3 páginasAmazing Facts - Exercises 1Alina DorobatAún no hay calificaciones
- Cell Division GizmoDocumento3 páginasCell Division Gizmoapi-522847737Aún no hay calificaciones
- 2010 FulltextDocumento410 páginas2010 FulltextNajam Din SahitoAún no hay calificaciones