También podría gustarte
- The Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeDe EverandThe Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeCalificación: 4 de 5 estrellas4/5 (5794)
- The Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreDe EverandThe Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreCalificación: 4 de 5 estrellas4/5 (1090)
- Never Split the Difference: Negotiating As If Your Life Depended On ItDe EverandNever Split the Difference: Negotiating As If Your Life Depended On ItCalificación: 4.5 de 5 estrellas4.5/5 (838)
- Hidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceDe EverandHidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceCalificación: 4 de 5 estrellas4/5 (895)
- Grit: The Power of Passion and PerseveranceDe EverandGrit: The Power of Passion and PerseveranceCalificación: 4 de 5 estrellas4/5 (588)
- Shoe Dog: A Memoir by the Creator of NikeDe EverandShoe Dog: A Memoir by the Creator of NikeCalificación: 4.5 de 5 estrellas4.5/5 (537)
- The Perks of Being a WallflowerDe EverandThe Perks of Being a WallflowerCalificación: 4.5 de 5 estrellas4.5/5 (2104)
- The Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersDe EverandThe Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersCalificación: 4.5 de 5 estrellas4.5/5 (344)
- Elon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureDe EverandElon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureCalificación: 4.5 de 5 estrellas4.5/5 (474)
- Her Body and Other Parties: StoriesDe EverandHer Body and Other Parties: StoriesCalificación: 4 de 5 estrellas4/5 (821)
- The Sympathizer: A Novel (Pulitzer Prize for Fiction)De EverandThe Sympathizer: A Novel (Pulitzer Prize for Fiction)Calificación: 4.5 de 5 estrellas4.5/5 (121)
- The Emperor of All Maladies: A Biography of CancerDe EverandThe Emperor of All Maladies: A Biography of CancerCalificación: 4.5 de 5 estrellas4.5/5 (271)
- The Little Book of Hygge: Danish Secrets to Happy LivingDe EverandThe Little Book of Hygge: Danish Secrets to Happy LivingCalificación: 3.5 de 5 estrellas3.5/5 (400)
- The World Is Flat 3.0: A Brief History of the Twenty-first CenturyDe EverandThe World Is Flat 3.0: A Brief History of the Twenty-first CenturyCalificación: 3.5 de 5 estrellas3.5/5 (2259)
- The Yellow House: A Memoir (2019 National Book Award Winner)De EverandThe Yellow House: A Memoir (2019 National Book Award Winner)Calificación: 4 de 5 estrellas4/5 (98)
- Devil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaDe EverandDevil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaCalificación: 4.5 de 5 estrellas4.5/5 (266)
- A Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryDe EverandA Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryCalificación: 3.5 de 5 estrellas3.5/5 (231)
- Team of Rivals: The Political Genius of Abraham LincolnDe EverandTeam of Rivals: The Political Genius of Abraham LincolnCalificación: 4.5 de 5 estrellas4.5/5 (234)
- On Fire: The (Burning) Case for a Green New DealDe EverandOn Fire: The (Burning) Case for a Green New DealCalificación: 4 de 5 estrellas4/5 (74)
- The Unwinding: An Inner History of the New AmericaDe EverandThe Unwinding: An Inner History of the New AmericaCalificación: 4 de 5 estrellas4/5 (45)
- Al Borg Laboratories PriceDocumento23 páginasAl Borg Laboratories PriceCheche Bureche0% (2)
- Rise of ISIS: A Threat We Can't IgnoreDe EverandRise of ISIS: A Threat We Can't IgnoreCalificación: 3.5 de 5 estrellas3.5/5 (137)
- Hematology Cell Morphology ChartDocumento2 páginasHematology Cell Morphology ChartMiaoAún no hay calificaciones
- NFKB3Documento9 páginasNFKB3Ariadna Zamora GuevaraAún no hay calificaciones
- Reviews: Micrornas in Myocardial InfarctionDocumento8 páginasReviews: Micrornas in Myocardial InfarctionAriadna Zamora GuevaraAún no hay calificaciones
- Nrcardio 2014 178Documento11 páginasNrcardio 2014 178Ariadna Zamora GuevaraAún no hay calificaciones
- Nrcardio 2015 38Documento14 páginasNrcardio 2015 38Ariadna Zamora GuevaraAún no hay calificaciones
- Reviews: Noncoding Rnas and Myocardial FibrosisDocumento9 páginasReviews: Noncoding Rnas and Myocardial FibrosisAriadna Zamora GuevaraAún no hay calificaciones
- Nej Mic M 1511213Documento1 páginaNej Mic M 1511213Ariadna Zamora GuevaraAún no hay calificaciones
- Reviews: Long Noncoding Rnas in Cardiac Development and AgeingDocumento11 páginasReviews: Long Noncoding Rnas in Cardiac Development and AgeingAriadna Zamora GuevaraAún no hay calificaciones
- GraficasDocumento5 páginasGraficasAriadna Zamora GuevaraAún no hay calificaciones
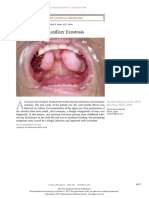
- Oral Maxillary Exostosis: Images in Clinical MedicineDocumento1 páginaOral Maxillary Exostosis: Images in Clinical MedicineAriadna Zamora GuevaraAún no hay calificaciones
- Antisense RNA TherapyDocumento17 páginasAntisense RNA TherapyGuleRia ShiVani100% (1)
- The Lymphatic System Aj ReportDocumento22 páginasThe Lymphatic System Aj ReportJohn Kirby DulayAún no hay calificaciones
- National Guide 3rd Adult Chart Sept 2018 WebDocumento2 páginasNational Guide 3rd Adult Chart Sept 2018 WebJenny NguyenAún no hay calificaciones
- Stras Chapter 10-12Documento17 páginasStras Chapter 10-12acer14appleAún no hay calificaciones
- Special Veterinary Pathology: Bacterial Diseases of Domestic AnimalsDocumento31 páginasSpecial Veterinary Pathology: Bacterial Diseases of Domestic AnimalsAnnie JohnAún no hay calificaciones
- A 1 Micro Globulin AdDocumento24 páginasA 1 Micro Globulin AdCARLOSAún no hay calificaciones
- EndocrineDocumento2 páginasEndocrineHoney Mie MorenoAún no hay calificaciones
- Intrinsically Disordered ProteinsDocumento30 páginasIntrinsically Disordered ProteinsJolanRuanAún no hay calificaciones
- MUST To KNOW in Immunohematology Blood BankingDocumento18 páginasMUST To KNOW in Immunohematology Blood BankingDanielJeremy DelaCruz Paragas79% (14)
- Biosci 1 200Documento22 páginasBiosci 1 200ROSENDA CORRALAún no hay calificaciones
- Practice Questions Section-A (2 Marks)Documento12 páginasPractice Questions Section-A (2 Marks)Preeti AgrawalAún no hay calificaciones
- CHN 215 - PTB CasepresDocumento57 páginasCHN 215 - PTB CasepresJeanneva Osorio EscoteAún no hay calificaciones
- BT CottonDocumento8 páginasBT Cottoniqra buttAún no hay calificaciones
- General Guidelines For The Disposal of CarcassesDocumento18 páginasGeneral Guidelines For The Disposal of CarcassesSuraj_SubediAún no hay calificaciones
- Kawasaki DiseaseDocumento14 páginasKawasaki Diseaseken100% (2)
- Genetic DisorderDocumento1 páginaGenetic DisorderKRISTELLE GEM CABANIGAún no hay calificaciones
- Decontamination Procedures TDocumento41 páginasDecontamination Procedures Tmochkurniawan100% (1)
- Malaria Research in South East AsiaDocumento186 páginasMalaria Research in South East AsiaRizal QowiAún no hay calificaciones
- Cell InjuryDocumento66 páginasCell InjuryHusam Abu DawoudAún no hay calificaciones
- Equine DiseasesDocumento65 páginasEquine Diseaseskreatura5Aún no hay calificaciones
- Lucid Medical Diagnostics PVT LTD (Banjara Hills) - HyderabadDocumento11 páginasLucid Medical Diagnostics PVT LTD (Banjara Hills) - HyderabadShashi KiranAún no hay calificaciones
- Genetic DiseasesDocumento28 páginasGenetic DiseasesdmorcfAún no hay calificaciones
- Reverse Genetics of RNA Viruses Methods and ProtocolsDocumento274 páginasReverse Genetics of RNA Viruses Methods and ProtocolsItrux jonathan cisnerosAún no hay calificaciones
- Gizzard Proventriculus Health Marker Avinews Int Sept 22Documento11 páginasGizzard Proventriculus Health Marker Avinews Int Sept 22mohamed helmyAún no hay calificaciones
- Cyto 2.4Documento5 páginasCyto 2.4Medtech SoonAún no hay calificaciones
- 12 General Microbiology, VirusDocumento22 páginas12 General Microbiology, VirusChaitanya BakliwalAún no hay calificaciones
- 5 6300732772877075609Documento131 páginas5 6300732772877075609Manoranjan sahuAún no hay calificaciones
- Avocado Storage TemperatureDocumento9 páginasAvocado Storage TemperatureOswald CastroAún no hay calificaciones