También podría gustarte
- Biodiversity: Explore the Diversity of Life on Earth with Environmental Science Activities for KidsDe EverandBiodiversity: Explore the Diversity of Life on Earth with Environmental Science Activities for KidsAún no hay calificaciones
- The Lives of Moths: A Natural History of Our Planet's Moth LifeDe EverandThe Lives of Moths: A Natural History of Our Planet's Moth LifeAún no hay calificaciones
- Chapter 2-1 Protozoa Diversity: Moss-Dwelling Micro-OrganismsDocumento16 páginasChapter 2-1 Protozoa Diversity: Moss-Dwelling Micro-OrganismsJose Facundo Ortega OrizAún no hay calificaciones
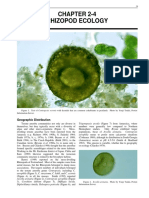
- Chapter 2-4 Rhizopod Ecology: Geographic DistributionDocumento8 páginasChapter 2-4 Rhizopod Ecology: Geographic DistributionJose Facundo Ortega OrizAún no hay calificaciones
- 5-1tardigrades Survival PDFDocumento20 páginas5-1tardigrades Survival PDFcemoaraAún no hay calificaciones
- 121 1995 Biodiversity at Its UtmostDocumento14 páginas121 1995 Biodiversity at Its UtmostArghya PaulAún no hay calificaciones
- Bryophytes: The Leap Onto LandDocumento27 páginasBryophytes: The Leap Onto LandaniruskAún no hay calificaciones
- SnailsDocumento8 páginasSnailsNicole De VegaAún no hay calificaciones
- Disney1994 Chapter EcologyDocumento32 páginasDisney1994 Chapter Ecologyairton.carvalhoAún no hay calificaciones
- Biodiversity HotspotDocumento2 páginasBiodiversity Hotspotr_marangga1432203Aún no hay calificaciones
- PoplarsandbiodiversityDocumento23 páginasPoplarsandbiodiversitySmit kalamkarAún no hay calificaciones
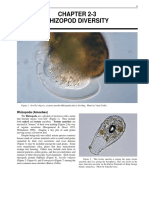
- Chapter 2-3 Rhizopod Diversity: Rhizopoda (Amoebas)Documento16 páginasChapter 2-3 Rhizopod Diversity: Rhizopoda (Amoebas)Jose Facundo Ortega OrizAún no hay calificaciones
- Fontaneto SyllabusDocumento11 páginasFontaneto SyllabusoscuridadAún no hay calificaciones
- Annelids TriviaDocumento2 páginasAnnelids TriviaRommel Urbano YabisAún no hay calificaciones
- Mites & Ticks33Documento19 páginasMites & Ticks33asaAún no hay calificaciones
- Mites & Ticks33Documento19 páginasMites & Ticks33asaAún no hay calificaciones
- Mites & Ticks33Documento19 páginasMites & Ticks33asaAún no hay calificaciones
- 1.2 The Microfauna: 1.2.1 ProtozoaDocumento19 páginas1.2 The Microfauna: 1.2.1 ProtozoaDidier tovarAún no hay calificaciones
- 1 TAW IntroDocumento151 páginas1 TAW IntroAlan MezaAún no hay calificaciones
- World Threatened AmphibiansDocumento625 páginasWorld Threatened AmphibiansThomasEngst100% (3)
- Geologic Time ScaleDocumento7 páginasGeologic Time ScaleWilliamAporboAún no hay calificaciones
- Mayaman Is ADocumento9 páginasMayaman Is ABayyina Bint BasharyAún no hay calificaciones
- Chapter 2-5 Peatland Rhizopods: Peatlands Taxa: SphagnumDocumento22 páginasChapter 2-5 Peatland Rhizopods: Peatlands Taxa: SphagnumJose Facundo Ortega OrizAún no hay calificaciones
- Spatial Distribution and Diversity of Small Commensal Mammals in Central Luzon State University and Its Helminth FaunaDocumento21 páginasSpatial Distribution and Diversity of Small Commensal Mammals in Central Luzon State University and Its Helminth FaunaElmer SulteroAún no hay calificaciones
- Living Things and The EnvironmentDocumento42 páginasLiving Things and The EnvironmentKavita RajaramAún no hay calificaciones
- Chapter 1 Aquatic Insects Why It Is Important To Deicated Our Time On Their StudyDocumento9 páginasChapter 1 Aquatic Insects Why It Is Important To Deicated Our Time On Their StudyIvan GonzalezAún no hay calificaciones
- Host Bromeliads and Their Associated Frog Species Furher Considerations On The Importance of Species Interactions For ConservationDocumento11 páginasHost Bromeliads and Their Associated Frog Species Furher Considerations On The Importance of Species Interactions For ConservationAna Maria MoncadaAún no hay calificaciones
- Definition Assignment English363Documento8 páginasDefinition Assignment English363api-644129074Aún no hay calificaciones
- Chapter 04 Joining ForcesDocumento6 páginasChapter 04 Joining ForcesEved1981 superrito.comAún no hay calificaciones
- Essential Entomology 2Nd Edition George C Mcgavin Full ChapterDocumento67 páginasEssential Entomology 2Nd Edition George C Mcgavin Full Chapterjoey.constanza138100% (4)
- Biology Assignment: Submitted To: Sir Khan Submitted By: Talha Rafiq Class: 8 Date of Submission: February 8, 2015Documento5 páginasBiology Assignment: Submitted To: Sir Khan Submitted By: Talha Rafiq Class: 8 Date of Submission: February 8, 2015Talha RafiqAún no hay calificaciones
- Provas de Proficiencia Ingles 1Documento10 páginasProvas de Proficiencia Ingles 1Daniel AguiarAún no hay calificaciones
- Essential EntomologyDocumento337 páginasEssential EntomologyHelio IshidaAún no hay calificaciones
- Fraser Et Al 2008Documento10 páginasFraser Et Al 2008kevin coquitoAún no hay calificaciones
- Wiki Loves MonumentsDocumento7 páginasWiki Loves MonumentsKvb SenabsolzurcAún no hay calificaciones
- Zoology BonDocumento118 páginasZoology BonRupkathar Pagol Kra RajkumariAún no hay calificaciones
- Gut Microbiome Analysis of Snails: A Biotechnological ApproachDocumento30 páginasGut Microbiome Analysis of Snails: A Biotechnological ApproachVladut BratfaleanAún no hay calificaciones
- Chapter 3 - InvertebratesDocumento28 páginasChapter 3 - InvertebratesJuan Carlos SánchezAún no hay calificaciones
- U1L4 - Keystone Species Critical ReadingDocumento4 páginasU1L4 - Keystone Species Critical Readingyohomieyougood123Aún no hay calificaciones
- Status of The Myxomycete Collection at The Uplb-Museum of Natural History (Uplb-Mnh) Mycological HerbariumDocumento15 páginasStatus of The Myxomycete Collection at The Uplb-Museum of Natural History (Uplb-Mnh) Mycological HerbariumCecilla Rosa PatiamAún no hay calificaciones
- Schill How To Culture Tardigrades FoodDocumento5 páginasSchill How To Culture Tardigrades FoodSant Iago Arc QuinAún no hay calificaciones
- Lecture 20 Notes 2013Documento4 páginasLecture 20 Notes 2013Richard HampsonAún no hay calificaciones
- Ants Are: (Video) Ants Gathering FoodDocumento24 páginasAnts Are: (Video) Ants Gathering FoodJordan MosesAún no hay calificaciones
- Water Beetles MalaysiaDocumento42 páginasWater Beetles MalaysiageobyAún no hay calificaciones
- What Is A Keystone Species WSDocumento3 páginasWhat Is A Keystone Species WSHusam AlSaifAún no hay calificaciones
- RagaeiOXDLHAVFKT75745 PDFDocumento6 páginasRagaeiOXDLHAVFKT75745 PDFMari DragomirAún no hay calificaciones
- Biodiversity and ConservationDocumento18 páginasBiodiversity and ConservationAsAún no hay calificaciones
- Book 1 - Unit 6Documento19 páginasBook 1 - Unit 6Phương NguyễnAún no hay calificaciones
- IntroductionDocumento21 páginasIntroductionsachinagsAún no hay calificaciones
- Feedinghabitsintrilobites: by andDocumento37 páginasFeedinghabitsintrilobites: by andkikiartAún no hay calificaciones
- Snail Is A: Slugs Semi-SlugsDocumento3 páginasSnail Is A: Slugs Semi-SlugsmmjpgcilAún no hay calificaciones
- An Initiatory Review On Bryophytes DiversityDocumento9 páginasAn Initiatory Review On Bryophytes DiversityIJRASETPublicationsAún no hay calificaciones
- Conduction in Bryophytes PDFDocumento8 páginasConduction in Bryophytes PDFRabindra Raj BistaAún no hay calificaciones
- Endemism AssignmentDocumento2 páginasEndemism AssignmentsherryAún no hay calificaciones
- Mandrillus Sphinx - Sexual Dimorphism and Physical Diversity of Individuals in Relation To Social StatusDocumento14 páginasMandrillus Sphinx - Sexual Dimorphism and Physical Diversity of Individuals in Relation To Social StatusJonathan Schwartz100% (2)
- Enriched Advanced Ecology Notes 2020Documento62 páginasEnriched Advanced Ecology Notes 2020aditi.ugandaAún no hay calificaciones
- Reticulitermes Flavipes: Geographic RangeDocumento8 páginasReticulitermes Flavipes: Geographic RangeHanifah HerliniAún no hay calificaciones
- Microbial Diversity: The Importance of Exploration and ConservationDocumento6 páginasMicrobial Diversity: The Importance of Exploration and ConservationRedd ZhuangAún no hay calificaciones
- Future of Life Reading GuideDocumento4 páginasFuture of Life Reading GuideJalen JonesAún no hay calificaciones
- Chapter 2-5 Peatland Rhizopods: Peatlands Taxa: SphagnumDocumento22 páginasChapter 2-5 Peatland Rhizopods: Peatlands Taxa: SphagnumJose Facundo Ortega OrizAún no hay calificaciones
- Chapter 2-3 Rhizopod Diversity: Rhizopoda (Amoebas)Documento16 páginasChapter 2-3 Rhizopod Diversity: Rhizopoda (Amoebas)Jose Facundo Ortega OrizAún no hay calificaciones
- Chapter 2-2 Protozoa: Ciliophora and Heliozoa Diversity: Other Ciliophora Known From BryophytesDocumento6 páginasChapter 2-2 Protozoa: Ciliophora and Heliozoa Diversity: Other Ciliophora Known From BryophytesJose Facundo Ortega OrizAún no hay calificaciones
- Hepaticas Foliosas de America Latina Memoirsofnewyork1119newyDocumento420 páginasHepaticas Foliosas de America Latina Memoirsofnewyork1119newyJose Facundo Ortega OrizAún no hay calificaciones
- 5A 5SN Talk On Young People Are ObeseDocumento9 páginas5A 5SN Talk On Young People Are ObesemasturinaAún no hay calificaciones
- Mohammad Ali LeghariDocumento64 páginasMohammad Ali LeghariNaoAún no hay calificaciones
- Article in Press: Neuroscience and Biobehavioral ReviewsDocumento10 páginasArticle in Press: Neuroscience and Biobehavioral ReviewsSahrul FajarAún no hay calificaciones
- 321 E Lesson 4Documento21 páginas321 E Lesson 4Novia eka PutriAún no hay calificaciones
- Anti Diabetic HerbsDocumento25 páginasAnti Diabetic HerbsSmritaPradhan PAún no hay calificaciones
- 501 Critical Reading QuestionsDocumento283 páginas501 Critical Reading QuestionsMenhaj Fatemioun100% (1)
- Pcod and InositolDocumento11 páginasPcod and Inositolkanishka upadhyayAún no hay calificaciones
- Dona Remedios Trinidad Romualdez Medical Foundation College of NursingDocumento3 páginasDona Remedios Trinidad Romualdez Medical Foundation College of NursingRoyce Vincent Tizon100% (3)
- Aims and Objectives of Cooking FoodDocumento6 páginasAims and Objectives of Cooking FoodShovan HazraAún no hay calificaciones
- Academic Writing Paragraph StructureDocumento2 páginasAcademic Writing Paragraph StructurechosunAún no hay calificaciones
- An Anti-Inflammatory Diet May Be Good For Your Joints - Harvard HealthDocumento10 páginasAn Anti-Inflammatory Diet May Be Good For Your Joints - Harvard HealthAdil ShaikAún no hay calificaciones
- The Cornflake DietDocumento8 páginasThe Cornflake DietSorindah MolinaAún no hay calificaciones
- Applebee's Nutritional InformationDocumento10 páginasApplebee's Nutritional InformationStanleyAún no hay calificaciones
- Hormonal Regulation of Farm Animal GrowthDocumento232 páginasHormonal Regulation of Farm Animal GrowthMihai MolnarAún no hay calificaciones
- SUPPLY SIDE West 2021Documento48 páginasSUPPLY SIDE West 2021Hebei Meishen SiraAún no hay calificaciones
- Effect of Essential Oil and Exogenous Enzymes On Blood Chemistry Profile and Antioxidant Capacity in Broiler ChickensDocumento5 páginasEffect of Essential Oil and Exogenous Enzymes On Blood Chemistry Profile and Antioxidant Capacity in Broiler ChickensEditor IJTSRDAún no hay calificaciones
- Discharge PlanDocumento2 páginasDischarge Plan2G Balatayo Joshua LeoAún no hay calificaciones
- COPD Case Study 1Documento15 páginasCOPD Case Study 1ifrah tariqAún no hay calificaciones
- BenfotiamineDocumento5 páginasBenfotiaminekether.thipharet5601Aún no hay calificaciones
- Lesson 4 Bricks 170Documento19 páginasLesson 4 Bricks 170Ninamae RomarateAún no hay calificaciones
- Health and Wellness Over The Counter ProductsDocumento19 páginasHealth and Wellness Over The Counter ProductsJorge Machado TencioAún no hay calificaciones
- Vitamin Deficiency Damaging Your BrainDocumento7 páginasVitamin Deficiency Damaging Your BrainMichael GemminkAún no hay calificaciones
- Pol Scie PPT FinalDocumento33 páginasPol Scie PPT FinalVenz LacreAún no hay calificaciones
- Guide To FitnessDocumento147 páginasGuide To FitnessLan MedinaAún no hay calificaciones
- The Definitive Guide To The "Push Pull Legs" Routine - Muscle For LifeDocumento25 páginasThe Definitive Guide To The "Push Pull Legs" Routine - Muscle For LifeMatt DerraughAún no hay calificaciones
- Treatment of Chronic Kidney DiseaseDocumento12 páginasTreatment of Chronic Kidney Diseaseaty100% (1)
- J. Biol. Chem.-1953-Forbes-359-66Documento9 páginasJ. Biol. Chem.-1953-Forbes-359-66Fachrun SofiyahAún no hay calificaciones
- ANS 312 - Applied Animal Nutrition Feedstuffs and Ration Formulation - OSU Extended Campus - Oregon State UniversityDocumento4 páginasANS 312 - Applied Animal Nutrition Feedstuffs and Ration Formulation - OSU Extended Campus - Oregon State UniversityRAHULAún no hay calificaciones
- Manufacturing Process of Refined SugarDocumento62 páginasManufacturing Process of Refined SugarVernie Soriano100% (1)
- Liver Cirrhosis Powerpoint (Week 2) REVISEDDocumento33 páginasLiver Cirrhosis Powerpoint (Week 2) REVISEDJustin CacheroAún no hay calificaciones