También podría gustarte
- Pink Floyd - Anthology (Song Book)Documento177 páginasPink Floyd - Anthology (Song Book)Porfírio Pinto100% (4)
- Herbal Medicine of The Republic of ChinaDocumento255 páginasHerbal Medicine of The Republic of ChinaarielighAún no hay calificaciones
- Introduction To Fourier Optics (J. Goodman 2nd SDocumento457 páginasIntroduction To Fourier Optics (J. Goodman 2nd Slinutez100% (3)
- Korean Cook BookDocumento64 páginasKorean Cook BookuvatsemotikeAún no hay calificaciones
- Ivan Galamian-Principles of Violin Playing and Teaching-Prentice Hall (1962) PDFDocumento66 páginasIvan Galamian-Principles of Violin Playing and Teaching-Prentice Hall (1962) PDFHéctor Abraham Olivas Namorado0% (5)
- Fields and Galois Theory: J.S. MilneDocumento138 páginasFields and Galois Theory: J.S. MilneAxelAún no hay calificaciones
- IMO 2015 ShorlistDocumento7 páginasIMO 2015 ShorlistAxelAún no hay calificaciones
- Homework1 SolDocumento4 páginasHomework1 SolAxelAún no hay calificaciones
- Cooperate Without Looking: Why We Care What People Think and Not Just What They DoDocumento6 páginasCooperate Without Looking: Why We Care What People Think and Not Just What They DoAxelAún no hay calificaciones
- Group Theory Via Rubik's CubeDocumento59 páginasGroup Theory Via Rubik's CubeYoshiaki Yonahasedo Oba100% (3)
- PNAS 2015 Chen 8002 7Documento6 páginasPNAS 2015 Chen 8002 7AxelAún no hay calificaciones
- 1964 EngDocumento1 página1964 EngChhorvorn VannAún no hay calificaciones
- Om Russian Math OlympiadDocumento21 páginasOm Russian Math Olympiadkoutsour27Aún no hay calificaciones
- The Yellow House: A Memoir (2019 National Book Award Winner)De EverandThe Yellow House: A Memoir (2019 National Book Award Winner)Calificación: 4 de 5 estrellas4/5 (98)
- Hidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceDe EverandHidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceCalificación: 4 de 5 estrellas4/5 (895)
- The Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeDe EverandThe Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeCalificación: 4 de 5 estrellas4/5 (5794)
- The Little Book of Hygge: Danish Secrets to Happy LivingDe EverandThe Little Book of Hygge: Danish Secrets to Happy LivingCalificación: 3.5 de 5 estrellas3.5/5 (399)
- Devil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaDe EverandDevil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaCalificación: 4.5 de 5 estrellas4.5/5 (266)
- Shoe Dog: A Memoir by the Creator of NikeDe EverandShoe Dog: A Memoir by the Creator of NikeCalificación: 4.5 de 5 estrellas4.5/5 (537)
- Elon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureDe EverandElon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureCalificación: 4.5 de 5 estrellas4.5/5 (474)
- Never Split the Difference: Negotiating As If Your Life Depended On ItDe EverandNever Split the Difference: Negotiating As If Your Life Depended On ItCalificación: 4.5 de 5 estrellas4.5/5 (838)
- Grit: The Power of Passion and PerseveranceDe EverandGrit: The Power of Passion and PerseveranceCalificación: 4 de 5 estrellas4/5 (588)
- A Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryDe EverandA Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryCalificación: 3.5 de 5 estrellas3.5/5 (231)
- The Emperor of All Maladies: A Biography of CancerDe EverandThe Emperor of All Maladies: A Biography of CancerCalificación: 4.5 de 5 estrellas4.5/5 (271)
- The World Is Flat 3.0: A Brief History of the Twenty-first CenturyDe EverandThe World Is Flat 3.0: A Brief History of the Twenty-first CenturyCalificación: 3.5 de 5 estrellas3.5/5 (2259)
- On Fire: The (Burning) Case for a Green New DealDe EverandOn Fire: The (Burning) Case for a Green New DealCalificación: 4 de 5 estrellas4/5 (73)
- The Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersDe EverandThe Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersCalificación: 4.5 de 5 estrellas4.5/5 (344)
- Rise of ISIS: A Threat We Can't IgnoreDe EverandRise of ISIS: A Threat We Can't IgnoreCalificación: 3.5 de 5 estrellas3.5/5 (137)
- Team of Rivals: The Political Genius of Abraham LincolnDe EverandTeam of Rivals: The Political Genius of Abraham LincolnCalificación: 4.5 de 5 estrellas4.5/5 (234)
- The Unwinding: An Inner History of the New AmericaDe EverandThe Unwinding: An Inner History of the New AmericaCalificación: 4 de 5 estrellas4/5 (45)
- The Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreDe EverandThe Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreCalificación: 4 de 5 estrellas4/5 (1090)
- The Sympathizer: A Novel (Pulitzer Prize for Fiction)De EverandThe Sympathizer: A Novel (Pulitzer Prize for Fiction)Calificación: 4.5 de 5 estrellas4.5/5 (121)
- The Perks of Being a WallflowerDe EverandThe Perks of Being a WallflowerCalificación: 4.5 de 5 estrellas4.5/5 (2102)
- Her Body and Other Parties: StoriesDe EverandHer Body and Other Parties: StoriesCalificación: 4 de 5 estrellas4/5 (821)
- Ipa18 202 SeDocumento15 páginasIpa18 202 SeDimas Suryo WicaksonoAún no hay calificaciones
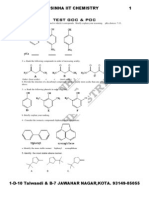
- Test1 Goc & Poc Tough by S.K.sinha See Chemistry Animations atDocumento3 páginasTest1 Goc & Poc Tough by S.K.sinha See Chemistry Animations atmyiitchemistry100% (1)
- The Municiplaity of Busia Solid Waste Management PlanDocumento15 páginasThe Municiplaity of Busia Solid Waste Management PlanToi JanetAún no hay calificaciones
- Complete Practical DistillingDocumento254 páginasComplete Practical DistillingMR X100% (5)
- 2014 Online Catlog - PDF MonDocumento50 páginas2014 Online Catlog - PDF Monjaag2000Aún no hay calificaciones
- SLR Strainer Data SheetDocumento7 páginasSLR Strainer Data SheetKailas NimbalkarAún no hay calificaciones
- Refrigeration Cycle, HVAC System Basics and Refrigerant Charging PDFDocumento13 páginasRefrigeration Cycle, HVAC System Basics and Refrigerant Charging PDFMurillo MendesAún no hay calificaciones
- Project Report On Pigment Print BinderDocumento4 páginasProject Report On Pigment Print BinderEIRI Board of Consultants and PublishersAún no hay calificaciones
- Hyperbaric Oxygen TherapyDocumento7 páginasHyperbaric Oxygen Therapy18juni1995Aún no hay calificaciones
- Plug Design Case StudyDocumento24 páginasPlug Design Case StudyAnonymous EsZwKlnBAún no hay calificaciones
- Eisco Industrial Catalogue - FinalDocumento276 páginasEisco Industrial Catalogue - FinalSOHAM AlhatAún no hay calificaciones
- Cleaning Validation MACO v2.1Documento3 páginasCleaning Validation MACO v2.1Syifa FatasyaaAún no hay calificaciones
- J.ultsonch.2014.08.022 InglesDocumento48 páginasJ.ultsonch.2014.08.022 InglesBrayan LuisAún no hay calificaciones
- Lampara de Fotocurado Bluephase MCDocumento72 páginasLampara de Fotocurado Bluephase MCErika PinillosAún no hay calificaciones
- Surgical Sutures & BandagesDocumento49 páginasSurgical Sutures & BandagesAnni Sholihah100% (1)
- TM 10-4930-220-13PDocumento133 páginasTM 10-4930-220-13PAdvocateAún no hay calificaciones
- 01-0019-01 Flame Methods Manual For AADocumento117 páginas01-0019-01 Flame Methods Manual For AARobert Larry Cordova DiazAún no hay calificaciones
- EagleBurgmann MG1 enDocumento4 páginasEagleBurgmann MG1 ensanjeevvangeAún no hay calificaciones
- LBR 2014 - ReDocumento62 páginasLBR 2014 - ReEugen XhaferajAún no hay calificaciones
- Drugdistributionfinal 151003021801 Lva1 App6891Documento12 páginasDrugdistributionfinal 151003021801 Lva1 App6891Raju NiraulaAún no hay calificaciones
- Fyup Chemistry SyllabusDocumento81 páginasFyup Chemistry SyllabusRaj KumarAún no hay calificaciones
- Born Oppenheimer ApproximationDocumento15 páginasBorn Oppenheimer ApproximationElizabeth HarrisonAún no hay calificaciones
- Redox EquilibriaDocumento2 páginasRedox Equilibriafunkykid80Aún no hay calificaciones
- Presenters Post16 Tcm18-118246Documento18 páginasPresenters Post16 Tcm18-118246Kamariah IsmailAún no hay calificaciones
- Paradip Refinery Project Write UpDocumento25 páginasParadip Refinery Project Write UpHarsha Jayaram100% (3)
- Orthodontic Wires - Properties / Orthodontic Courses by Indian Dental AcademyDocumento226 páginasOrthodontic Wires - Properties / Orthodontic Courses by Indian Dental Academyindian dental academy100% (1)
- Phenol SDocumento9 páginasPhenol SAnonymous 8rsxG4Aún no hay calificaciones
- Assignment 07 ADocumento3 páginasAssignment 07 AsundusAún no hay calificaciones
- Indian Regulations On Food PackgingDocumento20 páginasIndian Regulations On Food PackgingchoudhurykisanAún no hay calificaciones
- Embedded Enzymatic Biomaterial Degradation: 6836 Macromolecules 2009, 42, 6836-6839Documento4 páginasEmbedded Enzymatic Biomaterial Degradation: 6836 Macromolecules 2009, 42, 6836-6839hhkkllAún no hay calificaciones