También podría gustarte
- Absorción con filtro BK como terapia eficaz en el riñón del mieloma múltipleDe EverandAbsorción con filtro BK como terapia eficaz en el riñón del mieloma múltipleAún no hay calificaciones
- Enfermedades de Las PlaquetasDocumento3 páginasEnfermedades de Las PlaquetasBrayhan SánchezAún no hay calificaciones
- El Sistema InmuneDocumento15 páginasEl Sistema InmuneLeonela Marani NosedaAún no hay calificaciones
- Alteraciones de Las PlaquetasDocumento8 páginasAlteraciones de Las PlaquetasDaniela MuñozAún no hay calificaciones
- Ensayo NUEVE (Proteínas Totales Globulinas y Albúminas)Documento6 páginasEnsayo NUEVE (Proteínas Totales Globulinas y Albúminas)Cbz Sebastian Zermeño BrionesAún no hay calificaciones
- 20 Tipos de Proteínas y Sus Funciones en El CuerpoDocumento1 página20 Tipos de Proteínas y Sus Funciones en El CuerpoGuereca Montelongo Andrea100% (1)
- Plaquetas: estructura y funciónDocumento6 páginasPlaquetas: estructura y funciónmaria ramosAún no hay calificaciones
- Sindrome Nefrotico y NefriticoDocumento14 páginasSindrome Nefrotico y NefriticoMiguel Angel Mafla RoseroAún no hay calificaciones
- HematopoyesisDocumento13 páginasHematopoyesisLEO 999Aún no hay calificaciones
- Hemogram ADocumento36 páginasHemogram Asolorockerita100% (1)
- Histología de Las Glándulas Adrenales y El PáncreasfDocumento30 páginasHistología de Las Glándulas Adrenales y El PáncreasfCamila Rubilar TroncosoAún no hay calificaciones
- Tejido ConectivoDocumento27 páginasTejido ConectivoAle univalleAún no hay calificaciones
- Adaptación del sistema hepático: embriología y funcionesDocumento42 páginasAdaptación del sistema hepático: embriología y funcionesMaría Angelica DAppollonioAún no hay calificaciones
- Bases Moleculares de La Diapedesis PDFDocumento11 páginasBases Moleculares de La Diapedesis PDFRichard C. Munares100% (1)
- Enfermedad Renal Crónica - ApunteDocumento8 páginasEnfermedad Renal Crónica - ApunteAllison GómezAún no hay calificaciones
- Determinación del hematocritoDocumento10 páginasDeterminación del hematocritoAlejandraOzunaSolanoAún no hay calificaciones
- Importancia Del Seguimiento de Los Niveles Séricos de AntiepilépticosDocumento34 páginasImportancia Del Seguimiento de Los Niveles Séricos de AntiepilépticosChristian Escalante Delgado100% (1)
- Funcion Del Banco de SangreDocumento2 páginasFuncion Del Banco de SangreLeo GutierrezAún no hay calificaciones
- Anormalidades de EritrocitosDocumento58 páginasAnormalidades de EritrocitosLuis Uriel Gutierrez Diaz100% (1)
- Enfermedades ReumaticasDocumento21 páginasEnfermedades ReumaticasMarcelaDiazAún no hay calificaciones
- EspermiogramaDocumento6 páginasEspermiogramaHellen PlacidAún no hay calificaciones
- Pruebas de Diagnostico LeucemiasDocumento3 páginasPruebas de Diagnostico LeucemiasMyri RobalinoAún no hay calificaciones
- Dislipidemia ExposicionDocumento10 páginasDislipidemia ExposicionMonica Ospina100% (1)
- Fisiología plaquetaria: proceso de trombopoyesisDocumento4 páginasFisiología plaquetaria: proceso de trombopoyesisJosé Antonio Rodríguez PascualAún no hay calificaciones
- Formula LeucocitariaDocumento5 páginasFormula LeucocitariaJesus Loreto RangelAún no hay calificaciones
- Aterosclerosis y Lesión Endotelial.Documento7 páginasAterosclerosis y Lesión Endotelial.Luis Caspito GuevaraAún no hay calificaciones
- HemoglobinopatíasDocumento33 páginasHemoglobinopatíasMaría Teresa Bruno GAún no hay calificaciones
- Inflamacion Serosa y FibrinosaDocumento10 páginasInflamacion Serosa y FibrinosaRo Vizcaino SofiaAún no hay calificaciones
- Comunicación CelularDocumento8 páginasComunicación CelularJohn VelaAún no hay calificaciones
- Diatesis HemorragicaDocumento8 páginasDiatesis HemorragicaLuis SuarezAún no hay calificaciones
- VPHDocumento10 páginasVPHmonyckgcAún no hay calificaciones
- EriptosisDocumento2 páginasEriptosismi bella mas bellaAún no hay calificaciones
- Trombopoyesis y GranulopoyesisDocumento3 páginasTrombopoyesis y GranulopoyesisCarlosAún no hay calificaciones
- 1 - Tolerancia Autoinmunidad y TrasplanteDocumento13 páginas1 - Tolerancia Autoinmunidad y TrasplanteMEndar AlEeAún no hay calificaciones
- Sistema EndocrinoDocumento3 páginasSistema Endocrinomobufe2Aún no hay calificaciones
- Enzimas HepáticasDocumento24 páginasEnzimas HepáticasNohemy LopezAún no hay calificaciones
- Síntesis Del Grupo HemoDocumento9 páginasSíntesis Del Grupo HemoDario TaimalAún no hay calificaciones
- Péptido Natriurético AuricularDocumento3 páginasPéptido Natriurético Auricularivet_cabrera_1Aún no hay calificaciones
- ERITROPOYESISDocumento25 páginasERITROPOYESISgerman100% (1)
- Complicaciones PostransfusionalesDocumento25 páginasComplicaciones Postransfusionalesww.pacourgencias.blogspot.comAún no hay calificaciones
- Tincion GramDocumento6 páginasTincion GrameduardojmnzAún no hay calificaciones
- Fisiología plaquetaria y factores de la coagulación sanguíneaDocumento3 páginasFisiología plaquetaria y factores de la coagulación sanguíneaManu BGAún no hay calificaciones
- Revisión de los medicamentos hipolipemiantes más usados en la Región de MurciaDocumento8 páginasRevisión de los medicamentos hipolipemiantes más usados en la Región de MurciaAlejandro OlivardiaAún no hay calificaciones
- Liquido AmnioticDocumento49 páginasLiquido AmnioticJhon Andy Ramos100% (1)
- Epilepsia Mioclonica Con Fibras Rojas RasgadasDocumento26 páginasEpilepsia Mioclonica Con Fibras Rojas RasgadasLisseth SamudioAún no hay calificaciones
- Comunicación CelularDocumento49 páginasComunicación CelularRattus RattusAún no hay calificaciones
- Leucemia Mieloide Cronica.Documento15 páginasLeucemia Mieloide Cronica.Paulina Sanchez Sanchez100% (1)
- GlicopeptidosDocumento17 páginasGlicopeptidosapi-3750373100% (1)
- El Sistema de ComplementoDocumento22 páginasEl Sistema de ComplementoElianethDelAngel100% (1)
- Anormalidades LeucocitariasDocumento26 páginasAnormalidades LeucocitariasAlejandro Orozco CalderonAún no hay calificaciones
- Guia de Hematologia ClinicaDocumento27 páginasGuia de Hematologia ClinicaBetoLopezAún no hay calificaciones
- El Antígeno RPRDocumento5 páginasEl Antígeno RPRRafael ZevallosAún no hay calificaciones
- InmunologiaDocumento52 páginasInmunologiaMonica Mendoza100% (2)
- ART - Revision1 Chancroide-2014 PDFDocumento7 páginasART - Revision1 Chancroide-2014 PDFGAuReyesAún no hay calificaciones
- Sistema inmunitario: definición, componentes y funcionesDocumento17 páginasSistema inmunitario: definición, componentes y funcionesCarlos Zanns CarpioAún no hay calificaciones
- Cuadro Hematico CompletoDocumento19 páginasCuadro Hematico CompletoyancyAún no hay calificaciones
- Estructura y funciones de las inmunoglobulinasDocumento11 páginasEstructura y funciones de las inmunoglobulinasGloria Mardeni100% (1)
- Covid Fisiopatologia - En.esDocumento21 páginasCovid Fisiopatologia - En.eswiwiAún no hay calificaciones
- CrioglobulinasDocumento15 páginasCrioglobulinasglodovichiAún no hay calificaciones
- Manual para entender y tratar el asma: Consejos para mejorar la calidad de vidaDe EverandManual para entender y tratar el asma: Consejos para mejorar la calidad de vidaAún no hay calificaciones
- Papel Del EMG en Atrofia MonomielicaDocumento20 páginasPapel Del EMG en Atrofia MonomielicaVeronica MendozaAún no hay calificaciones
- Artefactos EEGDocumento4 páginasArtefactos EEGVeronica MendozaAún no hay calificaciones
- Prolapso ActualDocumento57 páginasProlapso ActualVeronica MendozaAún no hay calificaciones
- LaCANCION Del PuebloDocumento1 páginaLaCANCION Del PuebloVeronica MendozaAún no hay calificaciones
- Otro Dia Se VaDocumento2 páginasOtro Dia Se VaVeronica MendozaAún no hay calificaciones
- POCSDocumento9 páginasPOCSVeronica MendozaAún no hay calificaciones
- Tabla Resumen EMGDocumento1 páginaTabla Resumen EMGVeronica MendozaAún no hay calificaciones
- Chicas GuapasDocumento2 páginasChicas GuapasVeronica MendozaAún no hay calificaciones
- Encefalopatías en PediatriaDocumento20 páginasEncefalopatías en PediatriaVeronica MendozaAún no hay calificaciones
- Revision Eegvbmp02Documento20 páginasRevision Eegvbmp02Veronica MendozaAún no hay calificaciones
- Amo Del MesonDocumento4 páginasAmo Del MesonVeronica MendozaAún no hay calificaciones
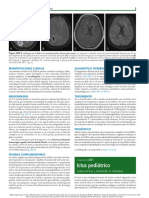
- Ictus PediátricoDocumento12 páginasIctus PediátricoVeronica MendozaAún no hay calificaciones
- Clasificacion ILAE 2017 Traducida 25 Jul 2018Documento54 páginasClasificacion ILAE 2017 Traducida 25 Jul 2018javier Uc08100% (2)
- EMG Músculos Esquema EASY EMGDocumento44 páginasEMG Músculos Esquema EASY EMGVeronica MendozaAún no hay calificaciones
- Protocolo CHUAC Muerte EncefalicaDocumento11 páginasProtocolo CHUAC Muerte EncefalicaVeronica MendozaAún no hay calificaciones
- Endocarditis InfecciosaDocumento5 páginasEndocarditis InfecciosaRafaella Valverde VilchrrezAún no hay calificaciones
- Dialnet LosTrastornosDeAnsiedadEnElDSM5 4803018Documento8 páginasDialnet LosTrastornosDeAnsiedadEnElDSM5 4803018Israel MeraAún no hay calificaciones
- Exploracion NLDocumento13 páginasExploracion NLVeronica MendozaAún no hay calificaciones
- Patología Benigna de VesículaDocumento61 páginasPatología Benigna de VesículaVeronica Mendoza100% (1)
- MONÓLOGO ILSE - SpAwDocumento2 páginasMONÓLOGO ILSE - SpAwVeronica MendozaAún no hay calificaciones
- Encuesta MASCOTASDocumento1 páginaEncuesta MASCOTASVeronica MendozaAún no hay calificaciones
- Pruebas Selectivas 2017/18: GrupoDocumento81 páginasPruebas Selectivas 2017/18: GrupoAriadna Pedrido AlonsoAún no hay calificaciones
- Asfixia PrenatalDocumento11 páginasAsfixia PrenatalWillian Cristobal Herrera SangronaAún no hay calificaciones
- PULMONARDocumento6 páginasPULMONARVeronica MendozaAún no hay calificaciones
- PrevalenciaVIHWaraoDocumento21 páginasPrevalenciaVIHWaraoVeronica MendozaAún no hay calificaciones
- Hijo de Madre DiabeticaDocumento5 páginasHijo de Madre DiabeticaJosue RamosAún no hay calificaciones
- Chronic Vomiting and Recurrent Pneumonia in An AdolescentDocumento11 páginasChronic Vomiting and Recurrent Pneumonia in An AdolescentVeronica MendozaAún no hay calificaciones
- Modelo Examen Tipo MIR Comentado 2Documento83 páginasModelo Examen Tipo MIR Comentado 2Veronica MendozaAún no hay calificaciones
- Luxaciones Glenohumerales PostraumáticasDocumento59 páginasLuxaciones Glenohumerales PostraumáticasVeronica MendozaAún no hay calificaciones
- MIR.01.1718.08.COMDocumento85 páginasMIR.01.1718.08.COMCatherine Saenz SerranoAún no hay calificaciones
- Fobias Específicas PDFDocumento109 páginasFobias Específicas PDFPsychieAún no hay calificaciones
- Manual de Primeros AuxiliosDocumento12 páginasManual de Primeros AuxiliosRosa SanchezAún no hay calificaciones
- Programa Ampliado de Inmunizaciones (PAI)Documento37 páginasPrograma Ampliado de Inmunizaciones (PAI)sebastian__narvaezAún no hay calificaciones
- Guia de Manejo Pacientes Picc OnconowDocumento2 páginasGuia de Manejo Pacientes Picc OnconowConny SaldiviaAún no hay calificaciones
- Plantas para Las CataratasDocumento3 páginasPlantas para Las Cataratasapi-3697492100% (4)
- La MononucleosisDocumento5 páginasLa MononucleosisNayla VelazcoAún no hay calificaciones
- Esquema de Planificación Del Texto Académico ExpositivoDocumento5 páginasEsquema de Planificación Del Texto Académico ExpositivoHENRRY ABEL CJANAHUIRE FERNANDEZAún no hay calificaciones
- Los Problemas de Espalda y Articulaciones Son Un Camino Directo Hacia Una Discapacidad InevitableDocumento3 páginasLos Problemas de Espalda y Articulaciones Son Un Camino Directo Hacia Una Discapacidad InevitableFernando Fernández FarjeAún no hay calificaciones
- Catedra PosterDocumento2 páginasCatedra PosterKatya UriarteAún no hay calificaciones
- PROCESO DEL NACIMIENTO Y SUS ETAPASDocumento3 páginasPROCESO DEL NACIMIENTO Y SUS ETAPASBilly Castañeda UrbinaAún no hay calificaciones
- Semiótica DermatológicaDocumento133 páginasSemiótica DermatológicaDavid SilvaAún no hay calificaciones
- Presentación 1Documento8 páginasPresentación 1Azuley RodriguezAún no hay calificaciones
- 2018-Carta de Derechos RS Revision PDFDocumento264 páginas2018-Carta de Derechos RS Revision PDFEfraín TeránAún no hay calificaciones
- Guia de Practicas Actual-1Documento46 páginasGuia de Practicas Actual-1Flor Bizarro0% (1)
- Dolor pélvico crónico: enfoque multifactorialDocumento63 páginasDolor pélvico crónico: enfoque multifactorialgrekezAún no hay calificaciones
- Evolución Histórica y Fundamento de La FisioterapiaDocumento10 páginasEvolución Histórica y Fundamento de La FisioterapiaDIANANAVASAún no hay calificaciones
- Mapa Del Sistema LinfaticoDocumento2 páginasMapa Del Sistema Linfaticoleilith cordova cordova100% (1)
- Electrocard Iograma (ECG) : Presentado Por: Daniela CarbonellDocumento43 páginasElectrocard Iograma (ECG) : Presentado Por: Daniela CarbonellDaniela CarbonellAún no hay calificaciones
- Evaluación paciente mareos agudosDocumento26 páginasEvaluación paciente mareos agudosRocio BueroAún no hay calificaciones
- 6 Situación de Salud Nutricional en ChileDocumento19 páginas6 Situación de Salud Nutricional en ChilePaula Karime NazerAún no hay calificaciones
- Cuadro Comparativo Marlon Cerrato 2023Documento3 páginasCuadro Comparativo Marlon Cerrato 2023Marlon CerratoAún no hay calificaciones
- Proyecto Emmanuel Bloque 1 Esp.Documento7 páginasProyecto Emmanuel Bloque 1 Esp.Guadalupe Monserrath Nolasco AlvarezAún no hay calificaciones
- Que Es El Masaje ReductivoDocumento22 páginasQue Es El Masaje ReductivoJulio PinosAún no hay calificaciones
- Procesos Fisiológicos Asociados A La DigestiónDocumento4 páginasProcesos Fisiológicos Asociados A La Digestiónsusana hoyos agudeloAún no hay calificaciones
- SDR 2 RespiratorioDocumento71 páginasSDR 2 RespiratorioGiselle STAún no hay calificaciones
- MÓDULO 2 FORMACION HUMANA (2) Hecho 2Documento34 páginasMÓDULO 2 FORMACION HUMANA (2) Hecho 2Arturo SepulvedaAún no hay calificaciones
- Constancia Cronologica YulianaDocumento4 páginasConstancia Cronologica YulianaMariela De TeranAún no hay calificaciones
- Taller de ColposDocumento58 páginasTaller de ColposMar Gil0% (1)
- Trabajo de Investigacion Practica GinecologiaDocumento11 páginasTrabajo de Investigacion Practica Ginecologiamiguelabc.rcAún no hay calificaciones
- Grupo BombayDocumento7 páginasGrupo BombayYudi Esther CampoAún no hay calificaciones