También podría gustarte
- Rojas Soriano Raúl 1983 3. Delimitación Del Tema de InvestigaciónDocumento15 páginasRojas Soriano Raúl 1983 3. Delimitación Del Tema de InvestigaciónLu GaravanoAún no hay calificaciones
- Comunicación NeuronalDocumento38 páginasComunicación NeuronalAtlamaltaAún no hay calificaciones
- SistNervioDocumento31 páginasSistNervioAlejandra EspinozaAún no hay calificaciones
- Neuronas: definición, características y funcionesDocumento27 páginasNeuronas: definición, características y funcionesLia CanoAún no hay calificaciones
- Filogénesis Del Sistema NerviosoDocumento46 páginasFilogénesis Del Sistema NerviosoEduardo VargasAún no hay calificaciones
- Comunicacion Neuronal SinapsisDocumento11 páginasComunicacion Neuronal Sinapsisvictor_cordova_66100% (1)
- 3 Unidades Funcionales de LuriaDocumento13 páginas3 Unidades Funcionales de LuriaTatiana Giraldo CandamilAún no hay calificaciones
- Desarrollo Ontogenetico Del SNDocumento5 páginasDesarrollo Ontogenetico Del SNCony Carrasco Chavez100% (1)
- La HabituacionDocumento10 páginasLa HabituacionAntoniaAún no hay calificaciones
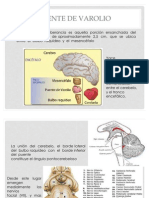
- Puente de VarolioDocumento10 páginasPuente de VarolioAndi VinsmokeAún no hay calificaciones
- Cuerpo calloso: anatomía y función de la principal comisura cerebralDocumento16 páginasCuerpo calloso: anatomía y función de la principal comisura cerebralEmmanuel GarciaAún no hay calificaciones
- Guía de Estudio Unidad 2 Motivación, Movimientos Forzados, Instinto e ImprontaDocumento3 páginasGuía de Estudio Unidad 2 Motivación, Movimientos Forzados, Instinto e ImprontaKuro Tengoku50% (2)
- Sentido Del TactoDocumento15 páginasSentido Del TactoJhon Flores100% (1)
- The Aversion ProyectDocumento7 páginasThe Aversion ProyectMqcUcr100% (1)
- Bases Neurobiologicas y Procesos AtencionalesDocumento4 páginasBases Neurobiologicas y Procesos AtencionalesAndres Felipe0% (1)
- NEURONADocumento3 páginasNEURONAleonardo david pinzónAún no hay calificaciones
- SNE-Organización y estructura del sistema nerviosoDocumento55 páginasSNE-Organización y estructura del sistema nerviosoClau HuertasAún no hay calificaciones
- Lóbulos CerebralesDocumento2 páginasLóbulos CerebralesDanny Bolaño MartinezAún no hay calificaciones
- Memoria DeclarativaDocumento6 páginasMemoria DeclarativaGABRIELA ANSELMIAún no hay calificaciones
- Cerebelo Equipo 4 Ayfi 2019Documento25 páginasCerebelo Equipo 4 Ayfi 2019Ruth Cobo RosalesAún no hay calificaciones
- Memoria y AprendizajeDocumento55 páginasMemoria y AprendizajeArleidy Tapie100% (1)
- Territorios Corticales de Proyección Motora, Sensitiva y Sensorial.Documento35 páginasTerritorios Corticales de Proyección Motora, Sensitiva y Sensorial.Luis Cristhian Urdiales Castillo100% (1)
- Células Nerviosas 2010Documento31 páginasCélulas Nerviosas 2010Camila Manzor100% (1)
- Celulas NerviosasDocumento10 páginasCelulas NerviosasRuben JuarezAún no hay calificaciones
- Depresión A Largo Plazo PDFDocumento4 páginasDepresión A Largo Plazo PDFMayra Herrera0% (1)
- Funcionamiento cerebral y psicopatologíaDocumento25 páginasFuncionamiento cerebral y psicopatologíaFrank Jhones100% (1)
- Teoria de Las Unidades Funcionales de LuriaDocumento37 páginasTeoria de Las Unidades Funcionales de LuriaLiliana MoraAún no hay calificaciones
- Alteraciones de la primera unidad funcional del cerebroDocumento1 páginaAlteraciones de la primera unidad funcional del cerebroRodrigo Montoya100% (1)
- Desarrollo Del Sistema NerviosoDocumento1 páginaDesarrollo Del Sistema NerviosoMily Figueroa100% (1)
- Plasticidad NeuronalDocumento3 páginasPlasticidad NeuronalElisa EspinozaAún no hay calificaciones
- Definición y Breve Historia de La NeuropsicologíaDocumento5 páginasDefinición y Breve Historia de La NeuropsicologíaDiana Rodriguez100% (1)
- Desarrollo Del Sistema NerviosoDocumento86 páginasDesarrollo Del Sistema NerviosoElena BallesterosAún no hay calificaciones
- Alexander LuriaDocumento21 páginasAlexander LuriaKaroll Astrid Silvestre Lagos100% (2)
- Modelos explicativos del proceso emocionalDocumento31 páginasModelos explicativos del proceso emocionalLaura Valentina Acosta RuizAún no hay calificaciones
- RataLab - Analizador Del Laberinto Circular de Barnes Basado en El Modelo de Mezclas GaussianasDocumento10 páginasRataLab - Analizador Del Laberinto Circular de Barnes Basado en El Modelo de Mezclas GaussianasAlejandroAún no hay calificaciones
- Ontogénesis Del Sistema NerviosoDocumento11 páginasOntogénesis Del Sistema NerviosoGabriela MendozaAún no hay calificaciones
- ReflejosDocumento6 páginasReflejosvictorAún no hay calificaciones
- Defectos Del Desarrollo de La LecturaDocumento5 páginasDefectos Del Desarrollo de La LecturaMiluskaSantosAún no hay calificaciones
- 2 Modelo Funcional Del Sistema NerviosoDocumento2 páginas2 Modelo Funcional Del Sistema NerviosoMicaela GoiteaAún no hay calificaciones
- Embriología Del Sistema Nervioso CentralDocumento20 páginasEmbriología Del Sistema Nervioso Centralkstar_17Aún no hay calificaciones
- Bases biológicas de la conducta humanaDocumento22 páginasBases biológicas de la conducta humanaJorge Rodríguez100% (1)
- SinapsisDocumento43 páginasSinapsisValeria EnrriquezAún no hay calificaciones
- Doctrina Neuronal de CajalDocumento2 páginasDoctrina Neuronal de CajalJavier Mohigefer BarreraAún no hay calificaciones
- Cerebro Conducta....... Enfoques Que Describen La Relacion Cerebro-ConductaDocumento7 páginasCerebro Conducta....... Enfoques Que Describen La Relacion Cerebro-ConductaJohanna Ponce0% (1)
- CEREBELODocumento9 páginasCEREBELOELIA ROSASAún no hay calificaciones
- La Importancia de Las Áreas de Percepción en El AprendizajeDocumento2 páginasLa Importancia de Las Áreas de Percepción en El Aprendizajeclaudia diaz100% (4)
- MARCO TEÓRICO - Los MillenialsDocumento14 páginasMARCO TEÓRICO - Los MillenialsVALENTINA FONTALVO LEYES ESTUDIANTEAún no hay calificaciones
- SISTEMA SENSITIVOMOTOR 2daDocumento25 páginasSISTEMA SENSITIVOMOTOR 2daPaola Garcia MejiaAún no hay calificaciones
- 1 Neuroanatomía Macroscópica ElementalDocumento17 páginas1 Neuroanatomía Macroscópica ElementalOscar Reynoso0% (1)
- La Memoria HumanaDocumento20 páginasLa Memoria Humanasiempreviva8480% (5)
- Historia y Naturaleza de La Psicologia Del DesarrolloDocumento5 páginasHistoria y Naturaleza de La Psicologia Del DesarrolloJhuliana VilchezAún no hay calificaciones
- Programa 1404 02 Prácticas de Psicobiología - P08 S-4Documento5 páginasPrograma 1404 02 Prácticas de Psicobiología - P08 S-4Manuel SánchezAún no hay calificaciones
- Bases Psicofisiológicas de Los Procesos Del Pensamiento y LenguajeDocumento11 páginasBases Psicofisiológicas de Los Procesos Del Pensamiento y LenguajeJoaquín C. Correa ICAún no hay calificaciones
- Las Tres Principales Unidades Funcionales de LuriaDocumento5 páginasLas Tres Principales Unidades Funcionales de LuriaRoxana Rojas0% (1)
- Cuerpo CallosoDocumento8 páginasCuerpo CallosoJavier CruzAún no hay calificaciones
- Sinapsis: comunicación neuronalDocumento9 páginasSinapsis: comunicación neuronalGaBu Alvarado100% (1)
- Lateralización hemisférica: Funciones cerebrales izquierda y derechaDocumento16 páginasLateralización hemisférica: Funciones cerebrales izquierda y derechaWilliam FryeAún no hay calificaciones
- Tronco CerebralDocumento4 páginasTronco CerebralAve FenixAún no hay calificaciones
- Las Neuronas y GliasDocumento10 páginasLas Neuronas y GliasDilania RodríguezAún no hay calificaciones
- Sistema Nervioso CienciasDocumento3 páginasSistema Nervioso CienciasBaezAlvAún no hay calificaciones
- Braunstein Nestor, Psiquiatría, Teoría Del Sujeto, Psicoanálisis (Hacia Lacan)Documento57 páginasBraunstein Nestor, Psiquiatría, Teoría Del Sujeto, Psicoanálisis (Hacia Lacan)David Téllez100% (3)
- Protozoarios Intestinales No Patógenos - AAMDocumento3 páginasProtozoarios Intestinales No Patógenos - AAMXiomara GarciaAún no hay calificaciones
- Universidad Nacional Daniel Alcides Carrion - Farmacos AntiparasitariosDocumento10 páginasUniversidad Nacional Daniel Alcides Carrion - Farmacos AntiparasitariosRicardo Perez-Saavedra CarruiteroAún no hay calificaciones
- Historia Clínica Congnitivo ConductualDocumento3 páginasHistoria Clínica Congnitivo ConductualFrancesca Idunno100% (1)
- Antibióticos: clasificación, mecanismos y usosDocumento10 páginasAntibióticos: clasificación, mecanismos y usosAna De Los Milagros Castillo Navarro100% (1)
- ACTIVIDAD - GUÍA #6 (1) de Lectura Critica Original90Documento9 páginasACTIVIDAD - GUÍA #6 (1) de Lectura Critica Original90Alejandro Rada PerezAún no hay calificaciones
- Serie Love at Stake 01 - Como Casarse Con Un Vampiro Millonario (Kerrelyn Sparks)Documento298 páginasSerie Love at Stake 01 - Como Casarse Con Un Vampiro Millonario (Kerrelyn Sparks)GuisBell100% (2)
- Protocolo quirúrgico COVID-19Documento14 páginasProtocolo quirúrgico COVID-19Elines GonzalezAún no hay calificaciones
- Test 4 - A - 2018Documento11 páginasTest 4 - A - 2018Raul MartinezAún no hay calificaciones
- Cuidado!.. Con El " Manzanillo"Documento5 páginasCuidado!.. Con El " Manzanillo"Rodrigo DominguezAún no hay calificaciones
- Epilepsia 292631 Downloadable 4399891Documento17 páginasEpilepsia 292631 Downloadable 4399891Keyla SernaqueAún no hay calificaciones
- Dialogo EkgDocumento6 páginasDialogo EkgDeisy CardenasAún no hay calificaciones
- Aislamientos hospitalarios: Estricto, Protector, por Gotas, Entérico y de ContactoDocumento2 páginasAislamientos hospitalarios: Estricto, Protector, por Gotas, Entérico y de ContactoAnellCarvajalAún no hay calificaciones
- Evaluacion Neuropsicologica en AutismoDocumento29 páginasEvaluacion Neuropsicologica en AutismoCaty Prieto100% (2)
- Funciones: OsteoclastoDocumento13 páginasFunciones: OsteoclastoClay MukurnereAún no hay calificaciones
- Adicción Al Móvil o Celular - FolletoDocumento2 páginasAdicción Al Móvil o Celular - FolletoEvelyn Karen Tito ChoqueAún no hay calificaciones
- Poder Telepatico PDFDocumento16 páginasPoder Telepatico PDFlector12100% (2)
- Efectos Del EtilenoDocumento4 páginasEfectos Del EtilenolunaAún no hay calificaciones
- Trastorno Conversivo y Crisis DisociativaDocumento3 páginasTrastorno Conversivo y Crisis DisociativaSoledadBella100% (1)
- Farmacodermias - Dra. TorreDocumento95 páginasFarmacodermias - Dra. TorreNatalia Loaiza ZamudioAún no hay calificaciones
- MetronidazolDocumento1 páginaMetronidazolBetssy Chavez AlbercaAún no hay calificaciones
- 02 - Relajantes de Musculo EstriadoDocumento45 páginas02 - Relajantes de Musculo EstriadowanopAún no hay calificaciones
- 827365SP L GenericsOnly v3Documento20 páginas827365SP L GenericsOnly v3LauraAún no hay calificaciones
- DSM5 Medidas Evaluacion Intensidad de Los Sintomas Dimensiones PsicóticasDocumento3 páginasDSM5 Medidas Evaluacion Intensidad de Los Sintomas Dimensiones Psicóticasロベルト ロベルトAún no hay calificaciones
- Vibraciones en Instalaciones y EdificiosDocumento32 páginasVibraciones en Instalaciones y EdificiosYules HernandezAún no hay calificaciones
- Funciones Pulmonares No RespiratoriasDocumento2 páginasFunciones Pulmonares No RespiratoriasMargarita del Carmen100% (1)
- Normas Seguridad LaboratorioDocumento20 páginasNormas Seguridad LaboratorioMENDOZA HUAMANI RAFAEL DANNYAún no hay calificaciones
- Nutricion EnteralDocumento18 páginasNutricion EnteralmollllyAún no hay calificaciones
- Recien Nacido ExpoDocumento24 páginasRecien Nacido ExpoisabellaAún no hay calificaciones
- Trabajo Final de Psicologia Del Desarrollo IIDocumento16 páginasTrabajo Final de Psicologia Del Desarrollo IInatividad morilloAún no hay calificaciones