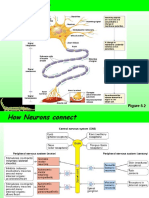
También podría gustarte
- Cellular and Molecular NeurophysiologyDe EverandCellular and Molecular NeurophysiologyCalificación: 5 de 5 estrellas5/5 (1)
- Surgical PhsyiologyDocumento170 páginasSurgical PhsyiologyTamjid Khan100% (2)
- Group 2 Azucena Bacud Baquiran Bautista - Neurophsyiology of Nerve Impulses Part 6-9Documento7 páginasGroup 2 Azucena Bacud Baquiran Bautista - Neurophsyiology of Nerve Impulses Part 6-9Timothy John BautistaAún no hay calificaciones
- Neurobiophysics2009 PreprintDocumento52 páginasNeurobiophysics2009 PreprintsmiglemAún no hay calificaciones
- EOG and its Applications in HealthcareDocumento48 páginasEOG and its Applications in HealthcareRose Edward50% (6)
- C-5 Signal TyransmissionDocumento14 páginasC-5 Signal TyransmissionSwad Abdul khalikAún no hay calificaciones
- Pharmacology of TheDocumento21 páginasPharmacology of TheKenneth NuñezAún no hay calificaciones
- Electrical Properties of BiomembranesDocumento52 páginasElectrical Properties of Biomembranesc3rberussAún no hay calificaciones
- Cellular Biology of EpileptogenesisDocumento10 páginasCellular Biology of EpileptogenesisandreacamachogonAún no hay calificaciones
- Advanced Cell BiologyDocumento14 páginasAdvanced Cell BiologyUmair AneesAún no hay calificaciones
- 2001 Intracellular Calcium and OuterDocumento6 páginas2001 Intracellular Calcium and OuterCaio LeônidasAún no hay calificaciones
- Latest Biochemical Techniques Capillary Electrophoresis: PrincipleDocumento9 páginasLatest Biochemical Techniques Capillary Electrophoresis: PrincipleVanessa SogieAún no hay calificaciones
- Wireless Electrical-Molecular Quantum Signalling For Cancer Cell ApoptosisDocumento18 páginasWireless Electrical-Molecular Quantum Signalling For Cancer Cell ApoptosisNoel Saúl Argüello SánchezAún no hay calificaciones
- Channelopathies: Ion Channel Defects Linked To Heritable Clinical DisordersDocumento12 páginasChannelopathies: Ion Channel Defects Linked To Heritable Clinical Disorderscote_cote6951Aún no hay calificaciones
- ManuscriptDocumento9 páginasManuscriptCélio MarquesAún no hay calificaciones
- Nervous System Electrical Signals and Action PotentialsDocumento20 páginasNervous System Electrical Signals and Action PotentialsAna G. VelascoAún no hay calificaciones
- Physio Lab Formal Report EXP 2 SEC-EDocumento7 páginasPhysio Lab Formal Report EXP 2 SEC-EVergel Jigs EnriquezAún no hay calificaciones
- SynapsesDocumento46 páginasSynapseshan abuur ahmedAún no hay calificaciones
- Electrophysiological Recording Techniques PDFDocumento7 páginasElectrophysiological Recording Techniques PDFAndrei TatomirAún no hay calificaciones
- What is electrophoresisDocumento22 páginasWhat is electrophoresismuskaan hameedAún no hay calificaciones
- Skeletal Muscle Physiology PDFDocumento6 páginasSkeletal Muscle Physiology PDFAstri Ggamjong Xiao LuAún no hay calificaciones
- Anggun Wening F - 1304617073 - Biolistrik Pada Sistem SarafDocumento8 páginasAnggun Wening F - 1304617073 - Biolistrik Pada Sistem SarafRosa AmaliaAún no hay calificaciones
- 111 Week 2 WorksheetDocumento4 páginas111 Week 2 WorksheetjabyleynesAún no hay calificaciones
- Biopsych Notes p2Documento28 páginasBiopsych Notes p2noorAún no hay calificaciones
- Numerical Study of The Electrothermal E Ffect On The Kinetic Reaction of Immunoassays For A Micro Uidic BiosensorDocumento8 páginasNumerical Study of The Electrothermal E Ffect On The Kinetic Reaction of Immunoassays For A Micro Uidic BiosensorMraidi SamehAún no hay calificaciones
- Nervous Coordination WorksheetDocumento4 páginasNervous Coordination WorksheetAshley BedassieAún no hay calificaciones
- Synapses HistologyDocumento6 páginasSynapses HistologySatwant SinghAún no hay calificaciones
- Physiology of Excitable Tissues LectureDocumento71 páginasPhysiology of Excitable Tissues LectureVikiAún no hay calificaciones
- Grupo 1 Practica 14Documento12 páginasGrupo 1 Practica 14Sadit Chauca VelaAún no hay calificaciones
- Surgical Physiology for MRCS Part 3Documento170 páginasSurgical Physiology for MRCS Part 3Steve FerisAún no hay calificaciones
- Photobiological Fundamentals of Low-Power Laser TherapyDocumento6 páginasPhotobiological Fundamentals of Low-Power Laser TherapyGabrielAún no hay calificaciones
- BiomecatronicaDocumento35 páginasBiomecatronicaAlan GrajedaAún no hay calificaciones
- 1.0 The Origin of Biopotentials (New)Documento62 páginas1.0 The Origin of Biopotentials (New)Hanif HussinAún no hay calificaciones
- Powerlab: Physiology of The Nerve: KeywordsDocumento7 páginasPowerlab: Physiology of The Nerve: KeywordsCJ RanesesAún no hay calificaciones
- P ('t':3) Var B Location Settimeout (Function (If (Typeof Window - Iframe 'Undefined') (B.href B.href ) ), 15000)Documento18 páginasP ('t':3) Var B Location Settimeout (Function (If (Typeof Window - Iframe 'Undefined') (B.href B.href ) ), 15000)hendra2darmawanAún no hay calificaciones
- Characteristics and Plasticity of Electrical SynapDocumento16 páginasCharacteristics and Plasticity of Electrical SynapjaneAún no hay calificaciones
- Measurement of Cellular Excitability by Whole Cell Patch Clamp TechniqueDocumento7 páginasMeasurement of Cellular Excitability by Whole Cell Patch Clamp TechniqueWai GeorgAún no hay calificaciones
- 1.17 Capillary Electrophoresis: The International Pharmacopoeia - Ninth Edition, 2019Documento8 páginas1.17 Capillary Electrophoresis: The International Pharmacopoeia - Ninth Edition, 2019asim awanAún no hay calificaciones
- Introduction To NeurophysiologyDocumento13 páginasIntroduction To Neurophysiologyvenkata ramakrishnaiahAún no hay calificaciones
- ElectrophysiologyDocumento32 páginasElectrophysiologyAbdelrehim Siraj100% (7)
- ElectrophysiologyDocumento32 páginasElectrophysiologyAbdelrehim SirajAún no hay calificaciones
- Resting Membrane PotentialDocumento22 páginasResting Membrane PotentialSanchezAún no hay calificaciones
- Neurophysiology: Biological membranes, synapses, neurotransmittersDocumento117 páginasNeurophysiology: Biological membranes, synapses, neurotransmittersRachel CajilesAún no hay calificaciones
- Electrical Resonance With Voltage-Gated Ion ChannelsDocumento8 páginasElectrical Resonance With Voltage-Gated Ion ChannelsNusrat JahanAún no hay calificaciones
- Molecular PathophysiologyDocumento3 páginasMolecular PathophysiologyElisa RoyAún no hay calificaciones
- CERS 2015 Poster SampleDocumento1 páginaCERS 2015 Poster SampleMichael WilsonAún no hay calificaciones
- Bio Potential and Bio ElectrodesDocumento39 páginasBio Potential and Bio ElectrodesSiva NareshAún no hay calificaciones
- Explain How Calcium Homeostasis Is Maintained, and Outline The Common Disorders of ThisDocumento3 páginasExplain How Calcium Homeostasis Is Maintained, and Outline The Common Disorders of ThisHani AlexAún no hay calificaciones
- Physiology Unit 4 - 231102 - 212048Documento23 páginasPhysiology Unit 4 - 231102 - 212048SanchezAún no hay calificaciones
- Chapter 48 Question Study online at quizlet.com/_3las3Documento3 páginasChapter 48 Question Study online at quizlet.com/_3las3Kalee Marion LemieuxAún no hay calificaciones
- Results: Michael A. Wilson, Timothy R. Brubaker, Andrea M. Mastro, Michael M. Micci, Sven G. Bilén, and Sean D. KnechtDocumento1 páginaResults: Michael A. Wilson, Timothy R. Brubaker, Andrea M. Mastro, Michael M. Micci, Sven G. Bilén, and Sean D. KnechtMichael WilsonAún no hay calificaciones
- Neuronal Functions and Electrical Signals in Under 40Documento59 páginasNeuronal Functions and Electrical Signals in Under 40sdsgdmfAún no hay calificaciones
- Smni83 Dynamics PDFDocumento32 páginasSmni83 Dynamics PDFLester IngberAún no hay calificaciones
- Lab 4Documento11 páginasLab 4api-27272391050% (2)
- Adams, P.R., Brown, D.A., and Constanti, A. (1982a) - M-Currents and Other Potassium Currents in Bullfrog Sympathetic Neurons.Documento36 páginasAdams, P.R., Brown, D.A., and Constanti, A. (1982a) - M-Currents and Other Potassium Currents in Bullfrog Sympathetic Neurons.Pavlos RigasAún no hay calificaciones
- Nerve and Muscle PhysiologyDocumento82 páginasNerve and Muscle PhysiologychandsriAún no hay calificaciones
- Neurochem FinalDocumento20 páginasNeurochem Finalelifulku.a1Aún no hay calificaciones
- Instrumentation Part 2Documento20 páginasInstrumentation Part 2Lawrence Genelago GamboaAún no hay calificaciones
- Electrophoresis and Capillary Electrophoresis PDFDocumento21 páginasElectrophoresis and Capillary Electrophoresis PDFVinay kumarAún no hay calificaciones
- Electro PermeablilizationDocumento7 páginasElectro Permeablilizationcindy tatiana rodriguez marinAún no hay calificaciones
- Wecb 467Documento31 páginasWecb 467qhqhqAún no hay calificaciones
- Wecb657 PDFDocumento10 páginasWecb657 PDFazzaassAún no hay calificaciones
- Wecb651 PDFDocumento15 páginasWecb651 PDFazzaassAún no hay calificaciones
- Wecb 424Documento12 páginasWecb 424azzaassAún no hay calificaciones
- Wecb451 PDFDocumento7 páginasWecb451 PDFazzaassAún no hay calificaciones
- Wecb435 PDFDocumento10 páginasWecb435 PDFazzaassAún no hay calificaciones
- Wecb641 PDFDocumento15 páginasWecb641 PDFazzaassAún no hay calificaciones
- Wecb657 PDFDocumento10 páginasWecb657 PDFazzaassAún no hay calificaciones
- Wecb451 PDFDocumento7 páginasWecb451 PDFazzaassAún no hay calificaciones
- Wecb641 PDFDocumento15 páginasWecb641 PDFazzaassAún no hay calificaciones
- Wecb 431Documento12 páginasWecb 431azzaassAún no hay calificaciones
- Wecb439 PDFDocumento12 páginasWecb439 PDFazzaassAún no hay calificaciones
- The Pentose Phosphate Pathway: An Overview: Advanced ArticleDocumento12 páginasThe Pentose Phosphate Pathway: An Overview: Advanced ArticleazzaassAún no hay calificaciones
- Wecb434 PDFDocumento13 páginasWecb434 PDFazzaassAún no hay calificaciones
- Wecb 424Documento12 páginasWecb 424azzaassAún no hay calificaciones
- Passive Diffusion Across Membranes: Advanced ArticleDocumento10 páginasPassive Diffusion Across Membranes: Advanced ArticleazzaassAún no hay calificaciones
- The Pentose Phosphate Pathway: An Overview: Advanced ArticleDocumento12 páginasThe Pentose Phosphate Pathway: An Overview: Advanced ArticleazzaassAún no hay calificaciones
- Wecb434 PDFDocumento13 páginasWecb434 PDFazzaassAún no hay calificaciones
- Passive Diffusion Across Membranes: Advanced ArticleDocumento10 páginasPassive Diffusion Across Membranes: Advanced ArticleazzaassAún no hay calificaciones
- Wecb 431Documento12 páginasWecb 431azzaassAún no hay calificaciones
- Wecb 424Documento12 páginasWecb 424azzaassAún no hay calificaciones
- Wecb 431Documento12 páginasWecb 431azzaassAún no hay calificaciones
- Wecb 417Documento10 páginasWecb 417azzaassAún no hay calificaciones
- Wecb 424Documento12 páginasWecb 424azzaassAún no hay calificaciones
- Wecb 423Documento7 páginasWecb 423azzaassAún no hay calificaciones
- Wecb 527Documento11 páginasWecb 527azzaassAún no hay calificaciones
- Wecb 423Documento7 páginasWecb 423azzaassAún no hay calificaciones
- Wecb 417Documento10 páginasWecb 417azzaassAún no hay calificaciones
- Wecb 688Documento14 páginasWecb 688azzaassAún no hay calificaciones
- Wecb 527Documento11 páginasWecb 527azzaassAún no hay calificaciones
- Biochemistry 4th Edition Voet Test Bank DownloadDocumento19 páginasBiochemistry 4th Edition Voet Test Bank Downloaddaddockstudderyxeq100% (24)
- General Physiology NursingDocumento113 páginasGeneral Physiology NursingYasuiAún no hay calificaciones
- Gabungan Laporan PraktikumDocumento21 páginasGabungan Laporan PraktikumHafizh ArrafiAún no hay calificaciones
- Nervous Systems - Neuron Function: Excitability Resting Potentials Graded Potential Action Potential The SynapseDocumento48 páginasNervous Systems - Neuron Function: Excitability Resting Potentials Graded Potential Action Potential The SynapseRajeev AgarwalAún no hay calificaciones
- PhysioEx Exercise 3-123Documento16 páginasPhysioEx Exercise 3-123NicoleAún no hay calificaciones
- John F. Butterworth, David C. Mackey, John D. Wasnick - Morgan & Mikhail's Clinical Anesthesiology-McGraw-Hill Education - Medical (2018)Documento21 páginasJohn F. Butterworth, David C. Mackey, John D. Wasnick - Morgan & Mikhail's Clinical Anesthesiology-McGraw-Hill Education - Medical (2018)fad02fadAún no hay calificaciones
- Muscular System Explained: Skeletal, Cardiac & Smooth Muscle TissueDocumento44 páginasMuscular System Explained: Skeletal, Cardiac & Smooth Muscle TissueClaudine Jo B. TalabocAún no hay calificaciones
- 8A The Nervous System and Neurones 8A CheckpointDocumento8 páginas8A The Nervous System and Neurones 8A CheckpointsalmaAún no hay calificaciones
- Neuro Complete ReviewDocumento138 páginasNeuro Complete ReviewMohsin FarooqAún no hay calificaciones
- Sensation and Perception 1st Edition Yantis Test BankDocumento15 páginasSensation and Perception 1st Edition Yantis Test Bankmelissaperezwzoeaksjgn100% (14)
- Neurons Notes 1 PDFDocumento16 páginasNeurons Notes 1 PDFabcdeAún no hay calificaciones
- NRSC500 Problem set: Key concepts in neurophysiologyDocumento7 páginasNRSC500 Problem set: Key concepts in neurophysiologyAnonymous grcOYV49fZAún no hay calificaciones
- 10 - Chapter - 01 Seizure DetectionDocumento21 páginas10 - Chapter - 01 Seizure DetectionSuguna NanthiniAún no hay calificaciones
- Anesthesia QuizDocumento4 páginasAnesthesia QuizMarites L. DomingoAún no hay calificaciones
- Physioex Lab Report: Pre-Lab Quiz ResultsDocumento4 páginasPhysioex Lab Report: Pre-Lab Quiz ResultsJoashan VacAún no hay calificaciones
- ECG & Cardiac ConductionDocumento78 páginasECG & Cardiac ConductionLalitaditya DivakarlaAún no hay calificaciones
- Computational Neuroscience - A First Course - 2013Documento142 páginasComputational Neuroscience - A First Course - 2013Sabahat Latif100% (1)
- Brain CureDocumento157 páginasBrain CureJosh BillAún no hay calificaciones
- Comparison Between Neuronal Action Potential and Action Potentials in Other CellsDocumento26 páginasComparison Between Neuronal Action Potential and Action Potentials in Other CellsmaryamAún no hay calificaciones
- 6 5 Nerves Hormones and HomeostasisDocumento19 páginas6 5 Nerves Hormones and Homeostasisapi-235355872Aún no hay calificaciones
- The Hodgkin-Huxley ModelDocumento7 páginasThe Hodgkin-Huxley ModelLike Hell I'm Telling YouAún no hay calificaciones
- (1.3) Soal UPM II Modul 1.3 2015Documento16 páginas(1.3) Soal UPM II Modul 1.3 2015isa infyAún no hay calificaciones
- BBMS3011 Synaptic Dysfunction and DepressionDocumento29 páginasBBMS3011 Synaptic Dysfunction and Depressionkeven319hk4304Aún no hay calificaciones
- Física Do Corpo Humano: Prof. Adriano Mesquita Alencar Dep. Física Geral Instituto de Física Da USPDocumento31 páginasFísica Do Corpo Humano: Prof. Adriano Mesquita Alencar Dep. Física Geral Instituto de Física Da USPMariaSuzanaDiazAún no hay calificaciones
- Anatomy and Physiology of Nervous SystemDocumento26 páginasAnatomy and Physiology of Nervous SystemDR. KUMARASWAMI HEALTH CENTRE COLLEGE OF NURSING KANYAKUMARIAún no hay calificaciones
- PL3102 NotesDocumento8 páginasPL3102 NotesKatriel HeartsBigbangAún no hay calificaciones
- How Pemf Therapy WorksDocumento33 páginasHow Pemf Therapy WorksDeepesh PariharAún no hay calificaciones
- Wbi15 01 Que 20240118Documento36 páginasWbi15 01 Que 20240118zainab zeinAún no hay calificaciones
- Pex 03 09Documento5 páginasPex 03 09Jila HafiziAún no hay calificaciones
- Chapter 48 Question Study online at quizlet.com/_3las3Documento3 páginasChapter 48 Question Study online at quizlet.com/_3las3Kalee Marion LemieuxAún no hay calificaciones
- Why We Die: The New Science of Aging and the Quest for ImmortalityDe EverandWhy We Die: The New Science of Aging and the Quest for ImmortalityCalificación: 3.5 de 5 estrellas3.5/5 (2)
- 10% Human: How Your Body's Microbes Hold the Key to Health and HappinessDe Everand10% Human: How Your Body's Microbes Hold the Key to Health and HappinessCalificación: 4 de 5 estrellas4/5 (33)
- The Molecule of More: How a Single Chemical in Your Brain Drives Love, Sex, and Creativity--and Will Determine the Fate of the Human RaceDe EverandThe Molecule of More: How a Single Chemical in Your Brain Drives Love, Sex, and Creativity--and Will Determine the Fate of the Human RaceCalificación: 4.5 de 5 estrellas4.5/5 (515)
- Masterminds: Genius, DNA, and the Quest to Rewrite LifeDe EverandMasterminds: Genius, DNA, and the Quest to Rewrite LifeAún no hay calificaciones
- The Consciousness Instinct: Unraveling the Mystery of How the Brain Makes the MindDe EverandThe Consciousness Instinct: Unraveling the Mystery of How the Brain Makes the MindCalificación: 4.5 de 5 estrellas4.5/5 (93)
- Crypt: Life, Death and Disease in the Middle Ages and BeyondDe EverandCrypt: Life, Death and Disease in the Middle Ages and BeyondCalificación: 4 de 5 estrellas4/5 (3)
- When the Body Says No by Gabor Maté: Key Takeaways, Summary & AnalysisDe EverandWhen the Body Says No by Gabor Maté: Key Takeaways, Summary & AnalysisCalificación: 3.5 de 5 estrellas3.5/5 (2)
- This Is Your Brain On Parasites: How Tiny Creatures Manipulate Our Behavior and Shape SocietyDe EverandThis Is Your Brain On Parasites: How Tiny Creatures Manipulate Our Behavior and Shape SocietyCalificación: 3.5 de 5 estrellas3.5/5 (31)
- The Ancestor's Tale: A Pilgrimage to the Dawn of EvolutionDe EverandThe Ancestor's Tale: A Pilgrimage to the Dawn of EvolutionCalificación: 4 de 5 estrellas4/5 (811)
- All That Remains: A Renowned Forensic Scientist on Death, Mortality, and Solving CrimesDe EverandAll That Remains: A Renowned Forensic Scientist on Death, Mortality, and Solving CrimesCalificación: 4.5 de 5 estrellas4.5/5 (397)
- The Other Side of Normal: How Biology Is Providing the Clues to Unlock the Secrets of Normal and Abnormal BehaviorDe EverandThe Other Side of Normal: How Biology Is Providing the Clues to Unlock the Secrets of Normal and Abnormal BehaviorAún no hay calificaciones
- Wayfinding: The Science and Mystery of How Humans Navigate the WorldDe EverandWayfinding: The Science and Mystery of How Humans Navigate the WorldCalificación: 4.5 de 5 estrellas4.5/5 (18)
- Undeniable: How Biology Confirms Our Intuition That Life Is DesignedDe EverandUndeniable: How Biology Confirms Our Intuition That Life Is DesignedCalificación: 4 de 5 estrellas4/5 (11)
- The Lives of Bees: The Untold Story of the Honey Bee in the WildDe EverandThe Lives of Bees: The Untold Story of the Honey Bee in the WildCalificación: 4.5 de 5 estrellas4.5/5 (44)
- The Second Brain: A Groundbreaking New Understanding of Nervous Disorders of the Stomach and IntestineDe EverandThe Second Brain: A Groundbreaking New Understanding of Nervous Disorders of the Stomach and IntestineCalificación: 4 de 5 estrellas4/5 (17)
- Eels: An Exploration, from New Zealand to the Sargasso, of the World's Most Mysterious FishDe EverandEels: An Exploration, from New Zealand to the Sargasso, of the World's Most Mysterious FishCalificación: 4 de 5 estrellas4/5 (30)
- Summary of Robert M. Sapolsky's DeterminedDe EverandSummary of Robert M. Sapolsky's DeterminedCalificación: 5 de 5 estrellas5/5 (2)
- A Brief History of Intelligence: Evolution, AI, and the Five Breakthroughs That Made Our BrainsDe EverandA Brief History of Intelligence: Evolution, AI, and the Five Breakthroughs That Made Our BrainsCalificación: 4.5 de 5 estrellas4.5/5 (4)
- Human Errors: A Panorama of Our Glitches, from Pointless Bones to Broken GenesDe EverandHuman Errors: A Panorama of Our Glitches, from Pointless Bones to Broken GenesCalificación: 3.5 de 5 estrellas3.5/5 (56)
- The Mind & The Brain: Neuroplasticity and the Power of Mental ForceDe EverandThe Mind & The Brain: Neuroplasticity and the Power of Mental ForceAún no hay calificaciones
- Gathering Moss: A Natural and Cultural History of MossesDe EverandGathering Moss: A Natural and Cultural History of MossesCalificación: 4.5 de 5 estrellas4.5/5 (347)
- The Dog Who Couldn't Stop Loving: How Dogs Have Captured Our Hearts for Thousands of YearsDe EverandThe Dog Who Couldn't Stop Loving: How Dogs Have Captured Our Hearts for Thousands of YearsAún no hay calificaciones
- Younger for Life: Feel Great and Look Your Best with the New Science of AutojuvenationDe EverandYounger for Life: Feel Great and Look Your Best with the New Science of AutojuvenationCalificación: 4 de 5 estrellas4/5 (1)
- Darwin's Dangerous Idea: Evolution and the Meaning of LifeDe EverandDarwin's Dangerous Idea: Evolution and the Meaning of LifeCalificación: 4 de 5 estrellas4/5 (523)
- Second Nature: A Gardener's EducationDe EverandSecond Nature: A Gardener's EducationCalificación: 4 de 5 estrellas4/5 (321)
- Superlative: The Biology of ExtremesDe EverandSuperlative: The Biology of ExtremesCalificación: 4.5 de 5 estrellas4.5/5 (51)
- Why We Sleep: Unlocking the Power of Sleep and DreamsDe EverandWhy We Sleep: Unlocking the Power of Sleep and DreamsCalificación: 4.5 de 5 estrellas4.5/5 (2083)
- Lymph & Longevity: The Untapped Secret to HealthDe EverandLymph & Longevity: The Untapped Secret to HealthCalificación: 4.5 de 5 estrellas4.5/5 (13)