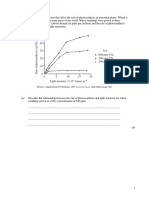
También podría gustarte
- Development of A Groundnut Core Collection Using Taxonomical, Geographical and Morphological DescriptorsDocumento10 páginasDevelopment of A Groundnut Core Collection Using Taxonomical, Geographical and Morphological DescriptorsJonahAún no hay calificaciones
- Application of Tissue Culture Techniques in PotatoDocumento18 páginasApplication of Tissue Culture Techniques in Potatokang_eebAún no hay calificaciones
- Modernization in Plant Breeding Approaches For Improving Biotic Stress Resistance in Crop PlantsDocumento16 páginasModernization in Plant Breeding Approaches For Improving Biotic Stress Resistance in Crop PlantsMuhammad Amir MaqboolAún no hay calificaciones
- Haploids in The Improvement of Crucifers (#160923) - 142283Documento6 páginasHaploids in The Improvement of Crucifers (#160923) - 142283Vikrant SahuAún no hay calificaciones
- Ehab ThesisDocumento133 páginasEhab Thesisehabmohamedzayed100% (2)
- Agronomy 11 01042Documento20 páginasAgronomy 11 01042Jose LuisAún no hay calificaciones
- tmpD928 TMPDocumento9 páginastmpD928 TMPFrontiersAún no hay calificaciones
- Invited Review Sugarcane Biotechnology: The Challenges and OpportunitiesDocumento19 páginasInvited Review Sugarcane Biotechnology: The Challenges and OpportunitiesGózalo LópezAún no hay calificaciones
- Regeneration and Transformation of An Elite Inbred Line of Maize (Zea Mays L.), With A Gene From Bacillus ThuringiensisDocumento7 páginasRegeneration and Transformation of An Elite Inbred Line of Maize (Zea Mays L.), With A Gene From Bacillus ThuringiensisCitlalli CedilloAún no hay calificaciones
- MicropropagationofBananaVarietiesMusaspp UsingShoot-TipCultureDocumento13 páginasMicropropagationofBananaVarietiesMusaspp UsingShoot-TipCultureVivek KawarkheAún no hay calificaciones
- EJDR - Volume 72 - Issue 1 - Pages 73-87Documento15 páginasEJDR - Volume 72 - Issue 1 - Pages 73-87Magali Ruiz RivasAún no hay calificaciones
- 760D84F31753Documento5 páginas760D84F31753Spam SpamAún no hay calificaciones
- TH 9769Documento80 páginasTH 9769Atul PharateAún no hay calificaciones
- Potato Cyst Nematodes: A New Threat To Potato Production in East AfricaDocumento13 páginasPotato Cyst Nematodes: A New Threat To Potato Production in East Africaabdneour zaouaguiAún no hay calificaciones
- KameswaraRao2017 Article AReviewOfFactorsThatInfluenceTDocumento14 páginasKameswaraRao2017 Article AReviewOfFactorsThatInfluenceTDunna DevisriAún no hay calificaciones
- Combining pearl millet disease resistanceDocumento6 páginasCombining pearl millet disease resistanceSri LakshmiAún no hay calificaciones
- Jee96 0914Documento11 páginasJee96 0914Aimer GuzmanAún no hay calificaciones
- Afr J BiotecDocumento11 páginasAfr J BiotecAugustine CrystalAún no hay calificaciones
- Rapid Culture of Pratylenchus Zeae On Carrot DiscsDocumento2 páginasRapid Culture of Pratylenchus Zeae On Carrot DiscsFrank KagodaAún no hay calificaciones
- Monoxenic Culture of Pratylenchus Zeae On Carrot DiscsDocumento2 páginasMonoxenic Culture of Pratylenchus Zeae On Carrot DiscsFernanda gAún no hay calificaciones
- Article1397576486 - Balogun Et AlDocumento9 páginasArticle1397576486 - Balogun Et AlFarah HabibahAún no hay calificaciones
- Ferrie 2011Documento9 páginasFerrie 2011László SágiAún no hay calificaciones
- Sabbavarapu2013 Article MolecularMappingOfQTLsForResis PDFDocumento13 páginasSabbavarapu2013 Article MolecularMappingOfQTLsForResis PDFMárioHenriqueAún no hay calificaciones
- Doubled Haploid Production in Capsicum Annuum L. Using Anther Culture: A ReviewDocumento6 páginasDoubled Haploid Production in Capsicum Annuum L. Using Anther Culture: A ReviewCarlos Rodríguez MartínAún no hay calificaciones
- Agronomy 02 00074Documento42 páginasAgronomy 02 00074ERYCK AZWARY ABRAHAM SURBAKTIAún no hay calificaciones
- Richa PlantsDocumento37 páginasRicha PlantsDr. Parmeshwar SahuAún no hay calificaciones
- Brassica Carinata Biology and Agronomy As A Biofuel CropDocumento18 páginasBrassica Carinata Biology and Agronomy As A Biofuel CropRamdeo SeepaulAún no hay calificaciones
- Morphological Characterization and Diversity Analysis of Adzuki Bean (Vigna Angularis (Willd.) Ohwi and Ohashi) GermplasmDocumento15 páginasMorphological Characterization and Diversity Analysis of Adzuki Bean (Vigna Angularis (Willd.) Ohwi and Ohashi) GermplasmAnjaliAún no hay calificaciones
- tmp43EC TMPDocumento7 páginastmp43EC TMPFrontiersAún no hay calificaciones
- Growth and Yield Performance of Pleurotus Ostreatus (Jacq. FR.) Kumm (Oyster Mushroom) On Different SubstratesDocumento7 páginasGrowth and Yield Performance of Pleurotus Ostreatus (Jacq. FR.) Kumm (Oyster Mushroom) On Different SubstratesSemAyat GoremsAún no hay calificaciones
- AndyGWAS-RevisedManuscript FileDocumento26 páginasAndyGWAS-RevisedManuscript FileZeleke WondimuAún no hay calificaciones
- Advances in chickpea breeding for increased yields and stress resistanceDocumento30 páginasAdvances in chickpea breeding for increased yields and stress resistancenarayananAún no hay calificaciones
- Field Crops Research: M. Oyekunle, B. Badu-Apraku, S. Hearne, J. FrancoDocumento11 páginasField Crops Research: M. Oyekunle, B. Badu-Apraku, S. Hearne, J. FrancoManuel GuzmanAún no hay calificaciones
- tmp374C TMPDocumento9 páginastmp374C TMPFrontiersAún no hay calificaciones
- Genetic Diversity and Conservation and Utilization of Plant Genetic ResourcesDocumento19 páginasGenetic Diversity and Conservation and Utilization of Plant Genetic ResourcesKaushal SurveAún no hay calificaciones
- NBT 2022 PDFDocumento10 páginasNBT 2022 PDFajaybioinfoAún no hay calificaciones
- Micropropagation of Banana Varieties (Musa SPP.) Using Shoot-Tip CultureDocumento12 páginasMicropropagation of Banana Varieties (Musa SPP.) Using Shoot-Tip Culturemaher Haji officialAún no hay calificaciones
- PigeonpeaKiran_8Documento30 páginasPigeonpeaKiran_8Adriano Dos SantosAún no hay calificaciones
- Feature: in Vitro Active vs. Field Genebank Maintenance of Sweet Potato Germplasm: Major Costs and ConsiderationsDocumento6 páginasFeature: in Vitro Active vs. Field Genebank Maintenance of Sweet Potato Germplasm: Major Costs and ConsiderationsrockfestAún no hay calificaciones
- Unguiculata L.) Due To Root-Knot Nematode, Meloidogyne Incognita Under Field ConditionsDocumento7 páginasUnguiculata L.) Due To Root-Knot Nematode, Meloidogyne Incognita Under Field ConditionsmucksoodganaieAún no hay calificaciones
- Micropropagation and conservation of chickpea genotypesDocumento9 páginasMicropropagation and conservation of chickpea genotypesbahiran alwaysAún no hay calificaciones
- Common Bean Genomics and Its Applications in Breeding ProgramsDocumento22 páginasCommon Bean Genomics and Its Applications in Breeding ProgramsrmbichiAún no hay calificaciones
- Diversity Analysis of Cotton (Gossypium Hirsutum L.) Germplasm Using The Cottonsnp63K ArrayDocumento20 páginasDiversity Analysis of Cotton (Gossypium Hirsutum L.) Germplasm Using The Cottonsnp63K ArrayJULIO CÉSAR CHÁVEZ GALARZAAún no hay calificaciones
- 834564v1 FullDocumento32 páginas834564v1 FullWoinshet MuhieAún no hay calificaciones
- A Run Kumare TalDocumento10 páginasA Run Kumare TalvitaAún no hay calificaciones
- In vitro Regeneration and Genetic Fidelity Assessment of Gladiolus hybridaDocumento10 páginasIn vitro Regeneration and Genetic Fidelity Assessment of Gladiolus hybridavitaAún no hay calificaciones
- Arun Kumar, Et AlDocumento10 páginasArun Kumar, Et AlvitaAún no hay calificaciones
- Ijab 2021Documento9 páginasIjab 2021EslamAún no hay calificaciones
- KuljarDocumento8 páginasKuljarRina Trihandayani PutriAún no hay calificaciones
- Genomic Resources in Plant Breeding For Sustainab - 2021 - Journal of Plant PhysDocumento18 páginasGenomic Resources in Plant Breeding For Sustainab - 2021 - Journal of Plant PhysTricia Marvi NavarroAún no hay calificaciones
- Productividas en Yuca y AsociadosDocumento20 páginasProductividas en Yuca y AsociadosjesusAún no hay calificaciones
- Evaluation of The Cross-Pollination in Maize (Zea Mays L.) Synthetic Varieties Grown in The High Guinean Savannah Zone ConditionsDocumento9 páginasEvaluation of The Cross-Pollination in Maize (Zea Mays L.) Synthetic Varieties Grown in The High Guinean Savannah Zone ConditionsMamta AgarwalAún no hay calificaciones
- Washburn 2019Documento15 páginasWashburn 2019KaniAún no hay calificaciones
- Development of yellow rust resistant wheat lines through wheat-maize crossesDocumento12 páginasDevelopment of yellow rust resistant wheat lines through wheat-maize crossesSwati KabraAún no hay calificaciones
- 2017 Current Status of Conventional and Molecular Interventions For Blast For Blast Resistance in RiceDocumento23 páginas2017 Current Status of Conventional and Molecular Interventions For Blast For Blast Resistance in RicekherapawanAún no hay calificaciones
- Common Bean as a Potential Crop for Future Food Security an Overview of Past Current and Future Contributions in Genomics Transcriptomics TransgenDocumento30 páginasCommon Bean as a Potential Crop for Future Food Security an Overview of Past Current and Future Contributions in Genomics Transcriptomics TransgenrmbichiAún no hay calificaciones
- Artigo 01 - Crop Biotechnology and The Future of FoodDocumento11 páginasArtigo 01 - Crop Biotechnology and The Future of FoodVinícius LopesAún no hay calificaciones
- Evaluation of Advanced Wheat (Triticum Aestivum L.) Lines For Stem Rust (Puccinia Graminis F. Sp. Tritici) Resistance and YieldDocumento14 páginasEvaluation of Advanced Wheat (Triticum Aestivum L.) Lines For Stem Rust (Puccinia Graminis F. Sp. Tritici) Resistance and YieldInternational Network For Natural SciencesAún no hay calificaciones
- Genetics: Diploid Chromosomes Polyploids Tetraploid Hexaploid EinkornDocumento2 páginasGenetics: Diploid Chromosomes Polyploids Tetraploid Hexaploid EinkornkoplakmanAún no hay calificaciones
- Genetic and Genomic Resources of Grain Legume ImprovementDe EverandGenetic and Genomic Resources of Grain Legume ImprovementAún no hay calificaciones
- F Distribution Significance LevelsDocumento9 páginasF Distribution Significance LevelskangpudjiAún no hay calificaciones
- ICARDADocumento2 páginasICARDADr-Muhammad Atif AttariAún no hay calificaciones
- F Distribution Significance LevelsDocumento9 páginasF Distribution Significance LevelskangpudjiAún no hay calificaciones
- Transfo of Tropical Grain Legumes Using Particle BombardmentDocumento1 páginaTransfo of Tropical Grain Legumes Using Particle BombardmentDr-Muhammad Atif AttariAún no hay calificaciones
- Transformation of Tropical Grain Legume Mungbean Using Particle BombardmentDocumento1 páginaTransformation of Tropical Grain Legume Mungbean Using Particle BombardmentDr-Muhammad Atif AttariAún no hay calificaciones
- Electroporation Protocol of Electrocompetent CellsDocumento4 páginasElectroporation Protocol of Electrocompetent CellsDr-Muhammad Atif AttariAún no hay calificaciones
- Transformation of Tropical Grain Legumes Using Particle BombardmentDocumento1 páginaTransformation of Tropical Grain Legumes Using Particle BombardmentDr-Muhammad Atif AttariAún no hay calificaciones
- Expressed Protein Ligation, A Novel Method For Studying Protein-Protein Interactions in TranscriptionDocumento5 páginasExpressed Protein Ligation, A Novel Method For Studying Protein-Protein Interactions in TranscriptionDr-Muhammad Atif AttariAún no hay calificaciones
- Efficient Transformation of Agrobacterium Spp. by High Voltage ElectroporationDocumento1 páginaEfficient Transformation of Agrobacterium Spp. by High Voltage ElectroporationDr-Muhammad Atif AttariAún no hay calificaciones
- A Proteomics Approach For Identifying Osmotic-Stress-Related Proteins in RiceDocumento12 páginasA Proteomics Approach For Identifying Osmotic-Stress-Related Proteins in RiceDr-Muhammad Atif AttariAún no hay calificaciones
- Agrobacterium Mediated Transformation of Chickpea (Cicer Arietinum L.) Embryo Axes PDFDocumento6 páginasAgrobacterium Mediated Transformation of Chickpea (Cicer Arietinum L.) Embryo Axes PDFDr-Muhammad Atif AttariAún no hay calificaciones
- Classification of Putative Transcription Factors of Medicago TruncatlaDocumento2 páginasClassification of Putative Transcription Factors of Medicago TruncatlaDr-Muhammad Atif AttariAún no hay calificaciones
- Bimolecular Fluorescence Complementation (BiFC) in Living Plant CellsDocumento3 páginasBimolecular Fluorescence Complementation (BiFC) in Living Plant CellsDr-Muhammad Atif AttariAún no hay calificaciones
- A Simple and Efficient Method For The Long-Term Preservation of Plant Cell Suspension Cultures PDFDocumento12 páginasA Simple and Efficient Method For The Long-Term Preservation of Plant Cell Suspension Cultures PDFDr-Muhammad Atif AttariAún no hay calificaciones
- Transformation of The Forage Legumes Medicago Sativa and OnobrychisDocumento2 páginasTransformation of The Forage Legumes Medicago Sativa and OnobrychisDr-Muhammad Atif AttariAún no hay calificaciones
- 25 Years of HIV ResearchDocumento26 páginas25 Years of HIV ResearchDr-Muhammad Atif AttariAún no hay calificaciones
- Eleven Golden Rules of RT PCRDocumento2 páginasEleven Golden Rules of RT PCRDr-Muhammad Atif AttariAún no hay calificaciones
- Antibiotic and Selection Reagent Usage For Resistant E. Coli CellsDocumento1 páginaAntibiotic and Selection Reagent Usage For Resistant E. Coli CellsDr-Muhammad Atif AttariAún no hay calificaciones
- 570 1Documento28 páginas570 1MASHAHZAD2004Aún no hay calificaciones
- National Education PolicyDocumento71 páginasNational Education PolicyDr-Muhammad Atif AttariAún no hay calificaciones
- Functional Analysis of Plant Cis ElementsDocumento50 páginasFunctional Analysis of Plant Cis ElementsDr-Muhammad Atif AttariAún no hay calificaciones
- ABFs, A Family of ABA-Responsive Element Binding FactorsDocumento8 páginasABFs, A Family of ABA-Responsive Element Binding FactorsDr-Muhammad Atif AttariAún no hay calificaciones
- Islamic Dua BookDocumento12 páginasIslamic Dua BookDr-Muhammad Atif AttariAún no hay calificaciones
- Diversity and Similarity Among Recognition Sequences of Dof Transcription FactorsDocumento6 páginasDiversity and Similarity Among Recognition Sequences of Dof Transcription FactorsDr-Muhammad Atif AttariAún no hay calificaciones
- European Council Teachers ConclusionsDocumento12 páginasEuropean Council Teachers ConclusionsDr-Muhammad Atif AttariAún no hay calificaciones
- The Dof Family of Plant Transcription FactorsDocumento6 páginasThe Dof Family of Plant Transcription FactorsDr-Muhammad Atif AttariAún no hay calificaciones
- فیصلہ کرنے کے مدنی پھول Faisla Karny ky madani PhoolDocumento58 páginasفیصلہ کرنے کے مدنی پھول Faisla Karny ky madani PhoolAshique Ali MughalAún no hay calificaciones
- Seed Composition The Physiology of Flowering PlantsDocumento1 páginaSeed Composition The Physiology of Flowering PlantsDr-Muhammad Atif AttariAún no hay calificaciones
- UAF Job Postings for Assistant ProfessorsDocumento3 páginasUAF Job Postings for Assistant ProfessorsDr-Muhammad Atif AttariAún no hay calificaciones
- Transgenics A New BreedDocumento3 páginasTransgenics A New BreedDr-Muhammad Atif AttariAún no hay calificaciones
- Biochemistry 8th Edition Berg Test BankDocumento36 páginasBiochemistry 8th Edition Berg Test Banklantautonomyurbeiu100% (38)
- Myank U.et Al. (1999) From Ethnomycology To Fungal BiotechnologyDocumento290 páginasMyank U.et Al. (1999) From Ethnomycology To Fungal BiotechnologyDániel KrüzselyiAún no hay calificaciones
- Herbal MedicineDocumento8 páginasHerbal MedicineStephanie Joy EscalaAún no hay calificaciones
- Slimming World RecipesDocumento73 páginasSlimming World Recipestelfordunited2100% (2)
- Ar PDFDocumento4 páginasAr PDFAriyansahAún no hay calificaciones
- HZS60 Concrete Batching Plant QuotationDocumento5 páginasHZS60 Concrete Batching Plant QuotationsherwinAún no hay calificaciones
- c4 and Cam PlantsDocumento34 páginasc4 and Cam PlantsOxman BhattiAún no hay calificaciones
- CHAPTER 4 BOTANY - Review Questions and AnswersDocumento7 páginasCHAPTER 4 BOTANY - Review Questions and AnswersPam GarciaAún no hay calificaciones
- THE DEAD RABBIT DRINKS MANUAL by Sean Muldoon and Jack McGarry PDFDocumento8 páginasTHE DEAD RABBIT DRINKS MANUAL by Sean Muldoon and Jack McGarry PDFAnderson LoveraAún no hay calificaciones
- True North Node 16Documento89 páginasTrue North Node 16Ynaffit Alteza Untal100% (1)
- Organic Cultivation of Medicinal PlantDocumento12 páginasOrganic Cultivation of Medicinal PlantVikrant PandeyAún no hay calificaciones
- Feg 2 Solved Assignment 2018 19 PDFDocumento10 páginasFeg 2 Solved Assignment 2018 19 PDFshivam sachanAún no hay calificaciones
- Soft LandscapeDocumento9 páginasSoft LandscapeAmal AmranAún no hay calificaciones
- Bill No. D1: External Works & Services (All Provisional) ITE MDocumento8 páginasBill No. D1: External Works & Services (All Provisional) ITE MRubbyRdAún no hay calificaciones
- Efikasi Tiga Jenis Rodentisida Antikoagulan Terhadap Hama Tikus Pada Perkebunan Kelapa SawitDocumento10 páginasEfikasi Tiga Jenis Rodentisida Antikoagulan Terhadap Hama Tikus Pada Perkebunan Kelapa Sawitpacul locoAún no hay calificaciones
- Cento Fine Foods - Fake San Marzano Tomatoes E.D.N.Y. - 2 - 19-cv-00974-JS-GRB - 14 - 0-14 - 14Documento217 páginasCento Fine Foods - Fake San Marzano Tomatoes E.D.N.Y. - 2 - 19-cv-00974-JS-GRB - 14 - 0-14 - 14Spencer SheehanAún no hay calificaciones
- No-Till Gardening: A Simple Method Using MulchDocumento7 páginasNo-Till Gardening: A Simple Method Using MulchShane Hodge100% (2)
- Intellectual Property Rights: Assistant Professor (Agril. Botany) RCSM College of Agriculture, KolhapurDocumento13 páginasIntellectual Property Rights: Assistant Professor (Agril. Botany) RCSM College of Agriculture, KolhapurJyoti IngoleAún no hay calificaciones
- Cholinesterase Inhibitors From Plants Po PDFDocumento17 páginasCholinesterase Inhibitors From Plants Po PDFVenkateswar RaoAún no hay calificaciones
- 4661 14945 2 PBDocumento6 páginas4661 14945 2 PBMaharani ZeinAún no hay calificaciones
- Research Paper 1Documento6 páginasResearch Paper 1api-302472440Aún no hay calificaciones
- Geographic Characteristics ChartDocumento3 páginasGeographic Characteristics Chartapi-295661927Aún no hay calificaciones
- Sandhaniya DasaimaniDocumento23 páginasSandhaniya DasaimanisnehalrajAún no hay calificaciones
- Label 2Documento2 páginasLabel 2Febiyanti SambiteAún no hay calificaciones
- Economic Survey Tables 2009-2010Documento132 páginasEconomic Survey Tables 2009-2010nikhilam.comAún no hay calificaciones
- Data Base Questions PhotosynthesisDocumento10 páginasData Base Questions PhotosynthesisValeCalderónAún no hay calificaciones
- Importance and Uses of Medicinal Plants - An Overview: December 2016Documento8 páginasImportance and Uses of Medicinal Plants - An Overview: December 2016samir sapdhareAún no hay calificaciones
- Grade: XI Subject: Commercial Fruit Production andDocumento2 páginasGrade: XI Subject: Commercial Fruit Production andNishad MallaAún no hay calificaciones
- Charles 2018 Tropcal Conservation ScienceDocumento5 páginasCharles 2018 Tropcal Conservation ScienceAlejandra LlerenaAún no hay calificaciones
- Amity Institute: Environmental StudiesDocumento12 páginasAmity Institute: Environmental StudiesArchanaGuptaAún no hay calificaciones