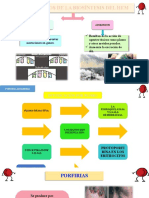
También podría gustarte
- Acceso a Universidad para Mayores de 25 años. Biología 2013-2017.: Solucionario Pruebas 2013-2017De EverandAcceso a Universidad para Mayores de 25 años. Biología 2013-2017.: Solucionario Pruebas 2013-2017Aún no hay calificaciones
- Metabolismo Del Hemo - SolDocumento6 páginasMetabolismo Del Hemo - SolOsvaldo HerraezAún no hay calificaciones
- 2 METABOLISMO de Las PorfirinasDocumento5 páginas2 METABOLISMO de Las PorfirinasEvelyn Guadalupe González BravoAún no hay calificaciones
- Porfirinas y Bilirrubinas - HarveyDocumento6 páginasPorfirinas y Bilirrubinas - HarveyJose MAún no hay calificaciones
- Porfirinas 2004Documento0 páginasPorfirinas 2004CesarAún no hay calificaciones
- Metabolismo HemeDocumento9 páginasMetabolismo Hemefialophora0210Aún no hay calificaciones
- Trabajo Final de Bioquímica II PorfiriaDocumento6 páginasTrabajo Final de Bioquímica II PorfiriaSofia BravoAún no hay calificaciones
- Secreción Biliar2Documento12 páginasSecreción Biliar2Maria Eduarda StevensAún no hay calificaciones
- Metabolismo de las porfirinas y sus defectos: las porfiriasDocumento5 páginasMetabolismo de las porfirinas y sus defectos: las porfiriasAgustin TevesAún no hay calificaciones
- Porfirinas y Pigmentos Biliares (Desgrabaci+ N)Documento7 páginasPorfirinas y Pigmentos Biliares (Desgrabaci+ N)Luisa LopezAún no hay calificaciones
- Metabolismo Grupo HEM (19-11)Documento5 páginasMetabolismo Grupo HEM (19-11)SDNKSJNDJKAún no hay calificaciones
- Sintesis y Degradacion Del HemoDocumento3 páginasSintesis y Degradacion Del HemoLeonel Carlos Rivero MamaniAún no hay calificaciones
- S23. Porfirinas y Pigmntos BiliaresDocumento28 páginasS23. Porfirinas y Pigmntos BiliaresGeraldine AnayaAún no hay calificaciones
- Porfirinas y Pigmentosbiliares (Diapositivas)Documento22 páginasPorfirinas y Pigmentosbiliares (Diapositivas)Jhusleidy España ArroyoAún no hay calificaciones
- PORFIRIA (Autoguardado) 2Documento16 páginasPORFIRIA (Autoguardado) 2Alejo ReyesAún no hay calificaciones
- Apuntes BQDocumento6 páginasApuntes BQJulissa Naomi Troya ToalaAún no hay calificaciones
- Metabolismo Del Grupo Hemo, Química y SíntesisDocumento26 páginasMetabolismo Del Grupo Hemo, Química y SíntesisBryan Pw SC TorresAún no hay calificaciones
- Porfirinas y Pigmentos BiliaresDocumento8 páginasPorfirinas y Pigmentos BiliaresPaola MoncadaAún no hay calificaciones
- Síntesis Del Grupo HemoDocumento9 páginasSíntesis Del Grupo HemoDario TaimalAún no hay calificaciones
- Grupo HemoDocumento20 páginasGrupo HemoDavid RecharteAún no hay calificaciones
- Sintesis Del Grupo Hemo 1Documento4 páginasSintesis Del Grupo Hemo 1tomasAún no hay calificaciones
- Metabolismo Del HemoDocumento3 páginasMetabolismo Del HemoMalena LaroccaAún no hay calificaciones
- Apuntes - Metabolismo de Las Porfirinas y Grupo HemoDocumento5 páginasApuntes - Metabolismo de Las Porfirinas y Grupo HemoLaura AybarAún no hay calificaciones
- Metabolismo Del Grupo HemoDocumento31 páginasMetabolismo Del Grupo HemoYaritza PinargtAún no hay calificaciones
- Catabolismo de Proteinas 2.3SPDocumento8 páginasCatabolismo de Proteinas 2.3SPDavid SanchezAún no hay calificaciones
- Las porfirinas: estructura, composición y aplicacionesDocumento55 páginasLas porfirinas: estructura, composición y aplicacionesMarín GEAún no hay calificaciones
- Proteínas 4Documento17 páginasProteínas 4Quique Villegas NavaAún no hay calificaciones
- Grupo Hemo y Porfirias.Documento32 páginasGrupo Hemo y Porfirias.Sabrina GaleanoAún no hay calificaciones
- Introducción al metabolismo del grupo hemoDocumento49 páginasIntroducción al metabolismo del grupo hemoRodrigo MullisacaAún no hay calificaciones
- Proteínas Plasmáticas e InmunoglobinasDocumento6 páginasProteínas Plasmáticas e InmunoglobinasAna FrancoAún no hay calificaciones
- Tarea Individual CalificadaDocumento8 páginasTarea Individual CalificadaYEYSON ELI SEGOVIA ZABALETAAún no hay calificaciones
- 4 Transtornos de La Biosíntesis Del HemDocumento13 páginas4 Transtornos de La Biosíntesis Del HemJael Fernandez FloresAún no hay calificaciones
- PorfirinasDocumento5 páginasPorfirinasAna Lilyam Rios BayonaAún no hay calificaciones
- Síntesis Del Grupo HemoDocumento4 páginasSíntesis Del Grupo HemoJorge Antonio Pérez Laines100% (1)
- Guia No 31Documento21 páginasGuia No 31Fernando HernandezAún no hay calificaciones
- Bioquimica en La SangreDocumento12 páginasBioquimica en La SangreMario Javier Ganoza MoralesAún no hay calificaciones
- Porfiria AnestesiaDocumento18 páginasPorfiria AnestesiaIrene Cristina100% (1)
- Metabolismo Del HemoDocumento46 páginasMetabolismo Del HemoJoham VegaAún no hay calificaciones
- Porfirinas y pigmentos biliares: importancia biomédicaDocumento34 páginasPorfirinas y pigmentos biliares: importancia biomédicaFrancisco GuerraAún no hay calificaciones
- Metabolismo de La HemoglobinaDocumento3 páginasMetabolismo de La HemoglobinaCharito Belizario ChecyaAún no hay calificaciones
- Dermatosis MetabólicasDocumento10 páginasDermatosis MetabólicasHel Es RoAún no hay calificaciones
- Anemia Sideroblastica TP 2Documento9 páginasAnemia Sideroblastica TP 2Mely MartínezAún no hay calificaciones
- Metabolismo de NucleótidosDocumento22 páginasMetabolismo de NucleótidosAngela gabriela Cometivos lescanAún no hay calificaciones
- Síntesis Del HemoDocumento8 páginasSíntesis Del HemoHenryk BazalarAún no hay calificaciones
- Hematologia. Síntesis y Degradación de Hemoglobina. UDELARDocumento39 páginasHematologia. Síntesis y Degradación de Hemoglobina. UDELARErika LoyarteAún no hay calificaciones
- Metabolismo fosfocálcico: regulación del calcio y fósforoDocumento12 páginasMetabolismo fosfocálcico: regulación del calcio y fósforoMaria Del Carmen Revi HerradAún no hay calificaciones
- Tema 9 BQ Clínica Y Patología MolecularDocumento13 páginasTema 9 BQ Clínica Y Patología MolecularMaría Karla Guerreiro GonzálezAún no hay calificaciones
- Bioquimica Médica. Aciduria OroticaDocumento10 páginasBioquimica Médica. Aciduria OroticaBeatriz A. Cázares33% (3)
- CLASE 4 - Amino Acdos, Peptidicos, ProteinaDocumento28 páginasCLASE 4 - Amino Acdos, Peptidicos, ProteinaAnonymous mAUbwzR2aBAún no hay calificaciones
- Metaolismo Grupo HemoDocumento35 páginasMetaolismo Grupo HemoClauMcClauAún no hay calificaciones
- Bilirrubina y Ácido ÚricoDocumento9 páginasBilirrubina y Ácido ÚricoL. Martin MorenoAún no hay calificaciones
- Porfirinas y Pigmentos BiliaresDocumento70 páginasPorfirinas y Pigmentos BiliaresJael Fernandez FloresAún no hay calificaciones
- Metabolismo de aminoácidos y proteínas: una guía concisaDocumento24 páginasMetabolismo de aminoácidos y proteínas: una guía concisaTeobaldoAún no hay calificaciones
- 2 Sintesis de PorfirinasDocumento6 páginas2 Sintesis de PorfirinasEvelyn Guadalupe González BravoAún no hay calificaciones
- Síntesis del grupo hemoDocumento5 páginasSíntesis del grupo hemoIngrid Alvarez AlvaradoAún no hay calificaciones
- Metabolismo de Nucleotidos.10Documento24 páginasMetabolismo de Nucleotidos.10Edwar Tarrillo Vasquez100% (1)
- BIOSINTESISDocumento8 páginasBIOSINTESISJubencio Anticona CuevaAún no hay calificaciones
- ANEMIA SeminarioDocumento8 páginasANEMIA SeminarioMARIA DLUCASAún no hay calificaciones
- Metabolismo de porfirinas en la UNJBG-TacnaDocumento41 páginasMetabolismo de porfirinas en la UNJBG-TacnaJesenia Fatima Paucar JoveAún no hay calificaciones
- Evaluación de La HemostasiaDocumento6 páginasEvaluación de La HemostasiaRUBY GALINDO SANDOVALAún no hay calificaciones
- TuberculosisDocumento3 páginasTuberculosisFREYER100% (1)
- Guía para La Elaboración Del Marco LógicoDocumento17 páginasGuía para La Elaboración Del Marco LógicoJairo Arosemena PerezAún no hay calificaciones
- Psicopatologia de La MemoriaDocumento19 páginasPsicopatologia de La MemoriaFREYERAún no hay calificaciones
- Ciclo de KrebsDocumento82 páginasCiclo de KrebsFREYERAún no hay calificaciones
- Población y Muestras en La InvestigaciónDocumento18 páginasPoblación y Muestras en La InvestigaciónFREYERAún no hay calificaciones
- Produccion Del CafeDocumento28 páginasProduccion Del CafeFREYER100% (1)
- Historia de El Derecho de Seguros en El PeruDocumento23 páginasHistoria de El Derecho de Seguros en El PeruFREYER100% (1)
- Emergencias ObstetricasDocumento20 páginasEmergencias ObstetricasFREYERAún no hay calificaciones
- El Virreinato Del PerúDocumento2 páginasEl Virreinato Del PerúFREYERAún no hay calificaciones
- RubeolaDocumento4 páginasRubeolaFREYERAún no hay calificaciones
- Como Hacer Un Proyecto PDFDocumento4 páginasComo Hacer Un Proyecto PDFFREYERAún no hay calificaciones
- Población y Muestras en La InvestigaciónDocumento18 páginasPoblación y Muestras en La InvestigaciónFREYERAún no hay calificaciones
- Control de Funciones VitalesDocumento9 páginasControl de Funciones VitalesFREYERAún no hay calificaciones
- Guia para La Administracion Segura de MedicamentosDocumento92 páginasGuia para La Administracion Segura de MedicamentosEfrén Atilano NolascoAún no hay calificaciones
- Primeros AuxiliosDocumento18 páginasPrimeros AuxiliosFREYERAún no hay calificaciones
- HemorragiasDocumento16 páginasHemorragiasFREYERAún no hay calificaciones
- Origen y Evolución Del HombreDocumento8 páginasOrigen y Evolución Del HombreFREYERAún no hay calificaciones
- 2ENZIMAS-I y IIDocumento39 páginas2ENZIMAS-I y IIFREYERAún no hay calificaciones
- Adaptación pulmonar a la altitudDocumento2 páginasAdaptación pulmonar a la altitudFREYERAún no hay calificaciones
- Glucolisis PDFDocumento3 páginasGlucolisis PDFEstefania Arizaga100% (1)
- METODOLOGIADocumento15 páginasMETODOLOGIAFREYERAún no hay calificaciones
- Valores de Referencia InmunoglobulinaDocumento5 páginasValores de Referencia InmunoglobulinaFREYERAún no hay calificaciones
- Biogénesis de RibosomasDocumento2 páginasBiogénesis de RibosomasFREYERAún no hay calificaciones
- Núcleo CelularDocumento11 páginasNúcleo CelularFREYERAún no hay calificaciones
- Manual HematologiaDocumento87 páginasManual Hematologiamiguel angel99% (192)
- Curso EKGDocumento54 páginasCurso EKGdra.hadley100% (7)
- La Aplicaciones A La MedicinaDocumento5 páginasLa Aplicaciones A La MedicinaFREYERAún no hay calificaciones
- II MacromoleculasDocumento42 páginasII MacromoleculasFREYERAún no hay calificaciones
- Hemo, Hierro y PorfiriasDocumento5 páginasHemo, Hierro y PorfiriasFREYERAún no hay calificaciones
- Primer Parcial Estomatologia 2019Documento3 páginasPrimer Parcial Estomatologia 2019Judith PalominoAún no hay calificaciones
- Cuál Es El Momento Indicado para TenerDocumento10 páginasCuál Es El Momento Indicado para TenerYeison castañoAún no hay calificaciones
- Dieta EquilibradaDocumento2 páginasDieta EquilibradaJose Diego PortugalAún no hay calificaciones
- Sedimento UrinarioDocumento10 páginasSedimento UrinarioEsmeraldaAún no hay calificaciones
- Percepción OlfativaDocumento15 páginasPercepción OlfativaFlamingAún no hay calificaciones
- Resveratrol PDFDocumento70 páginasResveratrol PDFMonicaAún no hay calificaciones
- Lesiones Nervios PerifericosDocumento56 páginasLesiones Nervios PerifericosStuart M.100% (1)
- CAPRINOMETRIADocumento5 páginasCAPRINOMETRIAEdson SaavedraAún no hay calificaciones
- Enfermedades de Transmisión SexualDocumento25 páginasEnfermedades de Transmisión SexualClaudia Suárez100% (1)
- Via IntradermicaDocumento22 páginasVia IntradermicaApril LP68% (19)
- El ácido alendrónico y su relación con la esofagitisDocumento29 páginasEl ácido alendrónico y su relación con la esofagitisaprolabgpAún no hay calificaciones
- Cirugia PulmonarDocumento10 páginasCirugia PulmonarErick Incio EsquivesAún no hay calificaciones
- INTRODUCCION-plexo BraquialDocumento7 páginasINTRODUCCION-plexo Braquialmayra894100% (1)
- ENFOQUE DEL MÉTODO DE BRUNNSTROM TotaDocumento73 páginasENFOQUE DEL MÉTODO DE BRUNNSTROM TotaGladis Pinto Mamani67% (6)
- Genética Médica para Médicos VeterinariosDocumento214 páginasGenética Médica para Médicos Veterinariosjm_garciap100% (1)
- Tumores Benignos de OvarioDocumento30 páginasTumores Benignos de OvarioMontse LuqueñoAún no hay calificaciones
- Anomalías Del PuerperioDocumento19 páginasAnomalías Del PuerperioDeisy Marcela Reyes ParraAún no hay calificaciones
- Ecologia de Los Sentidos ETOLOGIADocumento9 páginasEcologia de Los Sentidos ETOLOGIAMariaMarianaCortez0% (1)
- Trauma AbdominalDocumento33 páginasTrauma AbdominalAndrés Eduardo Reyes BelandriaAún no hay calificaciones
- Lectura #5Documento2 páginasLectura #5adri ramirezAún no hay calificaciones
- CircuncisiónDocumento29 páginasCircuncisiónGutiérrez CarlosAún no hay calificaciones
- Heridas MonografiasDocumento13 páginasHeridas MonografiasEveliaColalaClavoAún no hay calificaciones
- Enfermedades Digestivas en El AncianoDocumento28 páginasEnfermedades Digestivas en El AncianoTAnia Gordillo100% (1)
- Crónica de una adicción al fentanylDocumento13 páginasCrónica de una adicción al fentanylCarlos Bohorquez50% (2)
- Farmacos Hemostaticos y AnticoagulantesDocumento35 páginasFarmacos Hemostaticos y AnticoagulantesStephanie Alvarenga75% (4)
- Clase HTPDocumento21 páginasClase HTPapi-3703544100% (1)
- La Cueva TreintaDocumento125 páginasLa Cueva TreintaMelquíades Pérez PérezAún no hay calificaciones
- Informe Cientifico Sobre La Comunicación Materno-Filial en El Embarazo.Documento19 páginasInforme Cientifico Sobre La Comunicación Materno-Filial en El Embarazo.copyfree100% (5)
- Micosis Sistémicas o ProfundasDocumento28 páginasMicosis Sistémicas o ProfundasAna Yautibug0% (1)
- Meningitis Y EpilepsiaDocumento19 páginasMeningitis Y EpilepsiaAngela Mavarez0% (1)