También podría gustarte
- Shoe Dog: A Memoir by the Creator of NikeDe EverandShoe Dog: A Memoir by the Creator of NikeCalificación: 4.5 de 5 estrellas4.5/5 (537)
- The Yellow House: A Memoir (2019 National Book Award Winner)De EverandThe Yellow House: A Memoir (2019 National Book Award Winner)Calificación: 4 de 5 estrellas4/5 (98)
- Never Split the Difference: Negotiating As If Your Life Depended On ItDe EverandNever Split the Difference: Negotiating As If Your Life Depended On ItCalificación: 4.5 de 5 estrellas4.5/5 (838)
- Hidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceDe EverandHidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceCalificación: 4 de 5 estrellas4/5 (890)
- Grit: The Power of Passion and PerseveranceDe EverandGrit: The Power of Passion and PerseveranceCalificación: 4 de 5 estrellas4/5 (587)
- The Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeDe EverandThe Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeCalificación: 4 de 5 estrellas4/5 (5794)
- The Little Book of Hygge: Danish Secrets to Happy LivingDe EverandThe Little Book of Hygge: Danish Secrets to Happy LivingCalificación: 3.5 de 5 estrellas3.5/5 (399)
- Elon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureDe EverandElon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureCalificación: 4.5 de 5 estrellas4.5/5 (474)
- A Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryDe EverandA Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryCalificación: 3.5 de 5 estrellas3.5/5 (231)
- The Emperor of All Maladies: A Biography of CancerDe EverandThe Emperor of All Maladies: A Biography of CancerCalificación: 4.5 de 5 estrellas4.5/5 (271)
- The Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersDe EverandThe Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersCalificación: 4.5 de 5 estrellas4.5/5 (344)
- On Fire: The (Burning) Case for a Green New DealDe EverandOn Fire: The (Burning) Case for a Green New DealCalificación: 4 de 5 estrellas4/5 (73)
- Devil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaDe EverandDevil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaCalificación: 4.5 de 5 estrellas4.5/5 (265)
- The World Is Flat 3.0: A Brief History of the Twenty-first CenturyDe EverandThe World Is Flat 3.0: A Brief History of the Twenty-first CenturyCalificación: 3.5 de 5 estrellas3.5/5 (2219)
- Team of Rivals: The Political Genius of Abraham LincolnDe EverandTeam of Rivals: The Political Genius of Abraham LincolnCalificación: 4.5 de 5 estrellas4.5/5 (234)
- The Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreDe EverandThe Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreCalificación: 4 de 5 estrellas4/5 (1090)
- Rise of ISIS: A Threat We Can't IgnoreDe EverandRise of ISIS: A Threat We Can't IgnoreCalificación: 3.5 de 5 estrellas3.5/5 (137)
- The Unwinding: An Inner History of the New AmericaDe EverandThe Unwinding: An Inner History of the New AmericaCalificación: 4 de 5 estrellas4/5 (45)
- The Sympathizer: A Novel (Pulitzer Prize for Fiction)De EverandThe Sympathizer: A Novel (Pulitzer Prize for Fiction)Calificación: 4.5 de 5 estrellas4.5/5 (119)
- The Perks of Being a WallflowerDe EverandThe Perks of Being a WallflowerCalificación: 4.5 de 5 estrellas4.5/5 (2099)
- Her Body and Other Parties: StoriesDe EverandHer Body and Other Parties: StoriesCalificación: 4 de 5 estrellas4/5 (821)
- Krok 1 Stomatology: Test Items For Licensing ExaminationDocumento24 páginasKrok 1 Stomatology: Test Items For Licensing ExaminationhelloAún no hay calificaciones
- Cementoenamel Junction InsightsDocumento6 páginasCementoenamel Junction InsightsJessica ChenAún no hay calificaciones
- Handwritten Lympathic - Functions & Structures The Lymphatic System Transports Fluids To Blood - StudocuDocumento1 páginaHandwritten Lympathic - Functions & Structures The Lymphatic System Transports Fluids To Blood - StudocuAllyssa MarieAún no hay calificaciones
- Ayushman Spageric HerbalsDocumento6 páginasAyushman Spageric HerbalssubhalekhaAún no hay calificaciones
- Peritoneal SpaceDocumento54 páginasPeritoneal Spacehsah radiologyAún no hay calificaciones
- Orca Share Media1571800842460Documento27 páginasOrca Share Media1571800842460Reniel De MesaAún no hay calificaciones
- Accordion Sign-Appearance (C. Difficile)Documento41 páginasAccordion Sign-Appearance (C. Difficile)Andra HijratulAún no hay calificaciones
- Orthodontic Miniscrew Implants Clinical ApplicationsDocumento261 páginasOrthodontic Miniscrew Implants Clinical ApplicationsAbhishek ChauhanAún no hay calificaciones
- Digestive System Processes and Regulation - Anatomy and PhysiologyDocumento13 páginasDigestive System Processes and Regulation - Anatomy and PhysiologyFlonie DensingAún no hay calificaciones
- Lecture10 - Blood Groups and TransfusionDocumento28 páginasLecture10 - Blood Groups and Transfusionmohapatrarashmi050Aún no hay calificaciones
- Kundalini GlandDocumento4 páginasKundalini Glandvijender AtriAún no hay calificaciones
- An Essay On Yoga and HealthDocumento2 páginasAn Essay On Yoga and HealthGentleStrength100% (1)
- Regulation of Heart RateDocumento27 páginasRegulation of Heart RateGaurav SinghAún no hay calificaciones
- Insemination of Honey Bee Queens 4Documento32 páginasInsemination of Honey Bee Queens 4Novaaa RukmanaAún no hay calificaciones
- Otalgia & TinnitusDocumento19 páginasOtalgia & Tinnitushonda matizAún no hay calificaciones
- Three Circulatory Systems Working TogetherDocumento4 páginasThree Circulatory Systems Working TogetherSteven OwenAún no hay calificaciones
- ALLERGY AND IMMUNOLOGY OVERVIEWDocumento24 páginasALLERGY AND IMMUNOLOGY OVERVIEWmelAún no hay calificaciones
- Wiki Abdomen Mcqs ExplainedDocumento9 páginasWiki Abdomen Mcqs Explainedchemptnk100% (2)
- Thrombosis: Pgi Ricky G. JalecoDocumento34 páginasThrombosis: Pgi Ricky G. JalecoRicky JalecoAún no hay calificaciones
- PPT Bedah DigestifDocumento28 páginasPPT Bedah DigestifFatima Rima AndiniAún no hay calificaciones
- Linfedema: Actualización en El Diagnóstico y Tratamiento QuirúrgicoDocumento10 páginasLinfedema: Actualización en El Diagnóstico y Tratamiento QuirúrgicoHumberto Salas VelizAún no hay calificaciones
- Skeletal Maturity Indicators ReviewDocumento10 páginasSkeletal Maturity Indicators ReviewGina HyginusAún no hay calificaciones
- Pragyan College of Nursing-Bhopal BSC Nursing 1 Year Subject-Anatomy and Physiology Topic-Sense of Taste/TongueDocumento3 páginasPragyan College of Nursing-Bhopal BSC Nursing 1 Year Subject-Anatomy and Physiology Topic-Sense of Taste/TongueNeelofur Ibran AliAún no hay calificaciones
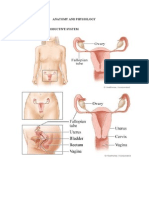
- The Female Reproductive Anatomy GuideDocumento10 páginasThe Female Reproductive Anatomy GuideBurni GuevaraAún no hay calificaciones
- Hydrocephalus: Vitya Chandika 2013-061-060 Priscila Stevanni 2013-061-066 Pricilia Nicholas 2013-061-070Documento24 páginasHydrocephalus: Vitya Chandika 2013-061-060 Priscila Stevanni 2013-061-066 Pricilia Nicholas 2013-061-070Priscila Stevanni100% (1)
- Mindmap Digestive AnatomyDocumento2 páginasMindmap Digestive Anatomyapi-535582646Aún no hay calificaciones
- Medical Surgical Nursing 1Documento43 páginasMedical Surgical Nursing 1Tifanny Shaine Tomas100% (2)
- IHbloodbank Midterm Topic 1Documento129 páginasIHbloodbank Midterm Topic 1Kat JornadalAún no hay calificaciones
- PediatricsDocumento33 páginasPediatricsnageshwarioshAún no hay calificaciones
- Encoding-Retrieval Similarity and MemoryDocumento48 páginasEncoding-Retrieval Similarity and Memorydsekulic_1Aún no hay calificaciones