Documentos de Académico
Documentos de Profesional
Documentos de Cultura
Motricidad Fina y Gruesa
Cargado por
MartitaVillalobosTítulo original
Derechos de autor
Formatos disponibles
Compartir este documento
Compartir o incrustar documentos
¿Le pareció útil este documento?
¿Este contenido es inapropiado?
Denunciar este documentoCopyright:
Formatos disponibles
Motricidad Fina y Gruesa
Cargado por
MartitaVillalobosCopyright:
Formatos disponibles
Motricidad fina y gruesa La Motricidad Fina La motricidad fina comprende todas aquellas actividades del nio que necesitan
de una precisin y un elevado nivel de coordinacin. Esta motricidad se refiere a los movimientos realizados por una o varias partes del cuerpo, que no tienen una amplitud sino que son movimientos de ms concrecin. Se cree que la motricidad fina se inicia hacia el ao y medio, cuando el nio, sin ningn aprendizaje y quizs movido por su instinto, empieza a poner objetos uno encima de otro, a hacer borrones con lpices, cambiar las cosas de sitio, etc.... Desarrollo de la Motricidad Fina EI desarrollo de la motricidad fina juega un papel central en el aumento de la inteligencia, debido a que se experimenta y aprende sobre su entorno. Las habilidades de motricidad fina se desarrollan en un orden progresivo, aunque se pueden dar grandes progresos y estancamientos o retrocesos sin consecuencias para el desarrollo normal del nio 0 a 12 meses: No hay control sobre las manos aunque al final del ao ya se nota el progreso. Si se toca su palma, cerrara su puo muy apretado, pero esto es una accin de reflejo inconsciente llamado "reflejo Darwinista", y desaparece en un plazo de dos a tres meses. As mismo, el infante agarrara un objeto puesto en su mano, pero sin ningn conocimiento de lo que esta haciendo. La coordinacin ojo-mano comienza a desarrollarse entre los 2 y 4 meses, comenzando as un periodo de prctica llamado ensayo y error al ver los objetos y tratar de tomarlos. A los cuatro o cinco meses, la mayora de los infantes pueden tomar un objeto que este dentro de su alcance, mirando solamente el objeto y no sus manos. Llamado "mximo nivel de alcance, este logro se considera un importante cimiento en el desarrollo de la motricidad fina. En el segundo semestre de esta etapa, comienzan a explorar y probar objetos antes de tomarlos. Uno de los logros motrices finos mas significativos es el tomar cosas usando los dedos como tenazas (pellizcado), lo cual aparece tpicamente entre las edades de 12 y 15 meses. 1 a 3 aos: Su desarrollo y curiosidad empujan al nio a manipular objetos cada vez de manera ms compleja, incluyendo la posibilidad de empujar palancas, girar las pginas de un libro, marcar nmeros del telfono, etc... Los dibujos que realizan son garabatos, pero empezaran a realizar figuras ms o menos circulares que les servirn de patrn para otros dibujos mas complejos. Jugarn con cubos y piezas que podrn poner una encima de otra hasta un cierto nivel. 3 a 5 aos (etapa pre-escolar): los retos en esta etapa, tales como el manejo de los cubiertos o atarse los zapatos, representan un salto evolutivo motriz importante. Cuando los nios tienen 3 aos, el control del lpiz puede ser grande y dibujan crculos sin que sean garabatos, animndose a dibujar figuras humanas o animales, aunque los trazos son muy simples. A los 4 aos, se usan las tijeras, se copian formas geomtricas y letras, se usan con criterio la plastilina y se pueden abrochar botones grandes. Algunos nios, usando la letra de palo, escriben su nombre y el de familiares o amigos cercanos. 5 aos (etapa escolar): A partir de aqu, la mayora de nios consolidan y avanzan claramente mas all del desarrollo logrado en la etapa pre-escolar, en sus habilidades motoras finas, perfeccionando lo adquirido. Pueden cortar, pegar y trazar formas con criterio. Pueden abrochar botones ms pequeos y tener control absoluto sobre aquellas tareas rutinarias, tanto en el mbito escolar como en el familiar. Los aspectos de la motricidad fina que se pueden trabajar a nivel escolar y educativo en general son: 1- Coordinacin Viso-Manual La coordinacin manual llevar a dominar su mano. Los elementos ms afectados, que intervienen directamente en este proceso son la mano, la mueca, el brazo y el antebrazo. Es muy importante tenerlo en cuenta ya que antes de que el nio pueda controlar sus movimientos en un espacio reducido como es un papel, ser necesario que pueda trabajar y dominar este gesto mas ampliamente en el suelo, pizarra y con elementos de poca precisin. Las actividades que ayudan a desarrollo la coordinacin visomanual son pintar, punzar, enhebrar, recortar, moldear, dibujar, colorear, etc... Foto: juego estimulante para la coordinacin y desarrollo motriz de manos y dedos 2- Coordinacin Facial Este es un aspecto de suma importancia ya que tiene dos componentes: 2.1- El dominio muscular 2.2- La posibilidad de comunicacin y relacin que tenemos con la gente que nos rodea a travs de nuestro cuerpo y especialmente de nuestros gestos voluntarios e involuntarios de la cara.
Es parte importante en la comunicacin del nio y se debe facilitar que controle y domine muscularmente su cara. Su importancia se debe a que permitir exteriorizar emociones, sentimientos y le ayudar a relacionarse, pues su cara dir con gestos lo que no sepa o pueda explicar con palabras 3- Coordinacin Fontica Se hace necesario su seguimiento y estimulacin, pues el nio deber emitir a lo largo de su madurez, la emisin sistemtica de cualquier sonido. Hay que estimular su desarrollo hablando, jugando e incitando al nio a relacionar objetos, personas, animales con sonidos, nombres, etc... Con ello ejercitaremos su parte fontica y a la vez, su memoria. La imitacin es un buen mtodo de aprendizaje. Hacia el ao y medio el nio puede tener la madurez para iniciar un lenguaje pero no contendr demasiadas palabras y las frases sern simples. Estos juegos motrices tendrn que continuar sobre todo para que el nio vaya adquiriendo un nivel de conciencia ms elevado. Entre los 2-3 aos el nio tiene posibilidades para sistematizar su lenguaje, para perfeccionar la emisin de sonidos y para concienciar la estructuracin de las frases y hacerlas cada vez ms complejas. Al final del tercer ao quedarn algunos sonidos para perfeccionar y unas irregularidades gramaticales y sintcticas a consolidar. Entre los 3 y 4 aos, consolidar y dominar el aparato fonador, hablar con total consciencia y coherencia y ser en la escuela donde madurar lingsticamente finalizando este proceso tan importante para la comunicacin. 4- Coordinacin gestual Se hace necesario el conocimiento de cada uno de los dedos individualmente y en conjunto para el dominio de las tareas, aunque no ser hasta los 10 aos cuando se asegurar su dominio. Hasta esa edad, las manos se ayudan (en la etapa pre-escolar) una a la otra en algunas tareas de precisin y ser el progreso escolar a partir de los 5 aos el que le llevar a la precisin y control individual de sus manos y dedos. La motricidad gruesa
Es la parte de la motricidad referente a los movimientos de los msculos que afectan a la locomocin o del desarrollo postural como andar, correr, saltar, etc., es decir, todo lo que tenga que ver con el desarrollo del nio que afectan a grupos de msculos sin tener en cuenta el detalle o la precisin que requiere la motricidad fina. A continuacin detallaremos el desarrollo de la motricidad gruesa en las diferentes fases de crecimiento. Fases de la motricidad gruesa. En el desarrollo motor, segn la edad, se observan cinco fases: 0 a 6 meses: Dependencia completa de la actividad refleja, en especial la succin. Hacia los tres o cuatro meses se inician los movimientos voluntarios debido a estmulos externos. 6 meses a 1 ao: Se caracteriza por la organizacin de las nuevas posibilidades del movimiento. Se observa una movilidad ms grande que se integra con la elaboracin del espacio y del tiempo. Esta organizacin sigue estrechamente ligada con la del tono muscular y la maduracin propia del proceso de crecimiento, la cul se enriquece continuamente debido al feedbak propio del desarrollo cognitivo. Cerca del ao, algunos nios caminan con ayuda. 1 a 2 aos: alrededor del ao y medio el nio anda solo y puede subir escalones con ayuda. Su curiosidad le hace tocar todo y se puede sentar en una silla, agacharse, etc..., A los 2 aos el nio corre y puede saltar con los dos pies juntos. Se pone en cuclillas, sube y baja las escaleras sintiendo el apoyo de la pared. Ejercicios de motricidad gruesa en etapa pre-escolar 3 a 4 aos: se consolida lo adquirido hasta el momento, se corre sin problemas, se suben y bajan escaleras sin ayuda ni apoyos, pueden ir de puntillas andando sobre las mismas sin problemas. Al llegar a los 4 aos se puede ir solo con un pie, el movimiento motor a lo largo del ao se ir perfeccionando hasta poder saltar, mover, subir y bajar por todas partes. 5 a 7 aos: El equilibrio entra en la fase determinante, donde se adquiere total autonoma en este sentido a lo largo de este perodo. En esta fase, se automatizan los conocimientos adquiridos hasta el momento, que sern la base de los nuevos conocimientos tanto internos como socio-afectivos. Las posibilidades que se abren al nio delante de las siguientes fases de crecimiento (adolescencia, pubertad) hasta llegar al desarrollo completo vendrn marcadas ineludiblemente por lo adquirido y consolidado en estas etapas. A partir de los 7 aos la maduracin est prcticamente completada, por lo que a partir de ahora y hasta los 12 aos es el momento idneo para realizar actividades que favorezcan el equilibrio y la coordinacin de movimientos.
Consecuencias de la carencia o deficiencia de vitamina A La carencia de vitamina A trae aparejado diversas consecuencias entra las que se destacan: Alteraciones oculares: puede ocasionar ceguera crepuscular, es decir disminuye la agudeza visual al anochecer, sensibilidad extrema a la luz como as tambin resecamiento, opacidad de la crnea con presencia de lceras, llamado xeroftalmia, la cual puede conducir a la ceguera Funciones de la vitamina A y el retinol en el organismo: sistema seo: es necesaria para el crecimiento y desarrollo de huesos. desarrollo celular: esencial para el crecimiento, mantenimiento y reparacin de las clulas de las mucosas, epitelios, piel, visin, uas, cabello y esmalte de dientes. sistema inmune: contribuye en la prevencin de enfermedades infecciosas, especialmente del aparato respiratorio creando barreras protectoras contra diferentes microorganismos. Estimula las funciones inmunes, entre ellas la respuesta de los anticuerpos y la actividad de varias clulas producidas por la medula sea que interviene en la defensa del organismo como fagocitos y linfocitos. Por ello promueve la reparacin de tejidos infectados y aumenta la resistencia a la infeccin. Sistema reproductivo: contribuye en la funcin normal de reproduccin, contribuyendo a la produccin de esperma como as tambin al ciclo normal reproductivo femenino. Debido a su rol vital en el desarrollo celular, la vitamina A ayuda a que los cambios que se producen en las clulas y tejidos durante el desarrollo del feto se desarrollen normalmente. Visin: es fundamental para la visin, ya que el Retinol contribuye a mejorar la visin nocturna, previniendo de ciertas alteraciones visuales como cataratas, glaucoma, perdida de visin, ceguera crepuscular ,tambin ayuda a combatir infecciones bacterianas como conjuntivitis. Antioxidante: previene el envejecimiento celular y la aparicin de cncer, ya que al ser un antioxidante natural elimina los radicales libres y protege al ADN de su accin mutagnica Volver al inicio de la pgina Las principales fuentes de vitamina A son: En el reino animal: los productos lcteos, la yema de huevo y el aceite de hgado de pescado. En los vegetales: En todos los vegetales amarillos a rojos, o verdes oscuros; zanahoria, batata, calabaza, zapallo, aj, espinacas, radiccio, lechuga, brcoli, coles de Bruselas, tomate, esprrago En las frutas: Damasco, durazno, meln, papaya, mango, mamn Acido Linoleico Conjugado (ALC) Lic. Marcela Licata
Es un cido graso esencial que lo produce la flora intestinal de los animales rumiantes, siempre a partir del cido linoleico. Esa produccin se ve aumentada cuando el animal es alimentado mediante el pastoreo y no cuando ingiere cereales. Se ve tambin en aceites vegetales (de maz, girasol, soja, etc.) aunque en una proporcin menor. Por lo tanto al ser producido por los rumiantes, la mejor fuente de C.L.A, son las carnes y los lcteos. Que sea un cido conjugado, significa que ha sufrido algn cambio en su estructura molecular, resultando ser nico y beneficioso para la salud y el bienestar general. Los seres humanos tambin producimos una pequea cantidad en el hgado, pero siempre a partir del cido linoleico. Sus efectos sobre el organismo Podemos decir que el C.L.A. es un nuevo nutriente que esta siendo utilizado para poder controlar el peso corporal. Est demostrado mediante diferentes estudios que el C.L.A. tiene diversas funciones:
reduce la grasa corporal,aumenta la masa muscular,remodela la figura corporal,reduce los niveles de colesterol y triglicridos en sangre,mejora el sistema inmune,acta como antioxidante, previniendo la aparicin de ciertas enfermedades y el envejecimiento prematuro, neutralizando los radicales libres,Para llegar a lograr los resultados anteriormente mencionados, siempre debe ser utilizado como complemento de una dieta hipocalrica y un plan de ejercicio fsico . La recomendacin es consumir 3 gramos diarios, pero con una dieta variada, lograremos alcanzar 1,5 gramos /da. Por consiguiente, si lo que se quiere es lograr resultados a nivel de reducir el peso corporal y la masa grasa, ser necesario tomar algn complemento que nos provea de C.L.A, siempre bajo supervisin de un profesional. Lo mismo sucede con las personas que tienen una alimentacin exenta de alimentos de origen animal, como por ejemplo los vegetarianos, donde la ingesta de C.LA est muy reducida. Pueden recurrir a suplementos para cubrir las carencias.Cul es el mecanismo por el cual se disminuye la grasa corporal? El C.L.A produce un bloqueo en el transporte de las grasas hacia las clulas adiposas. Esto se debe a que el C.L.A inhibe la funcin de la enzima LPL (lipoprotein lipasa) y al mismo tiempo favorece la liplisis (destruccin de la grasa). Ayuda a aumentar el transporte de grasa hacia la mitocondria celular (rgano de la clula encargado de la respiracin celular y produccin de energa) para ser oxidada.
Y cmo aumenta la masa muscular? El C.L.A aumenta la actividad de la enzima CPT (carnitina palmitoin transferasa). Esta enzima esta presente en el msculo y es la que transporta las grasas hacia la mitocondria para ser usada como combustible. La actividad fsica estimula entonces el transporte de la grasa hacia la clula muscular para ser oxidada. Por eso es que sin actividad fsica, el C.L.A no produce los mismos efectos.Dado el grave problema que representa en la actualidad la obesidad y el sobrepeso, este ingrediente natural ha cobrado gran importancia al combatir unos de los principales trastornos de este siglo. Cmo afectan las grasas de los alimentos sobre las grasas en nuestro cuerpo Una vez que sabemos qu tipo de grasas podemos encontrar en los alimentos (las grasas de los alimentos) y cmo estas se transforman y circulan por nuestro cuerpo convertidas en las diferentes fracciones lipdicas alimentos (las grasas en nuestro cuerpo), veremos como interactan las unas con las otras y de qu forma esto afecta al desarrollo de las enfermedades cardiovasculares.
Colesterol Acidos grasos saturados Acidos grasos monoinsaturados Acidos grasos poliinsaturados Acidos grasos trans Vitaminas antioxidantes
Colesterol A pesar del miedo que nos han hecho tenerle, el colesterol presente en los alimentos no es tan peligroso como el que circula por nuestras venas. En numerosos experimentos con diferentes especies de animales se encontr que el colesterol de la dieta resultaba ser altamente aterognico (formador de placas de ateroma en las arterias), por lo que se pens que en los humanos ocurrira lo mismo. Sin embargo, los humanos en general no son tan sensibles al colesterol de la dieta como otras especies de animales, y hoy en da tenemos la evidencia de que el colesterol ingerido influye bastante menos sobre el aumento de colesterol en sangre (que es el realmente peligroso) que el consumo de grasas saturadas. Esto se explica porque la absorcin del colesterol en el intestino humano est limitada a un 40 o 50 % de lo ingerido, con amplias diferencias de unos individuos a otros determinadas por factores genticos. Esta variabilidad tambin depende de numerosos factores. Por ejemplo, los triglicridos presentes en el intestino (de alimentos grasos) favorecen la absorcin de colesterol,
mientras que los esteroles vegetales (de alimentos ricos en fibra vegetal) y marinos (del marisco) la reducen por competir con su absorcin. El contenido de colesterol de la alimentacin tpica occidental es de unos 400 mg/da. Cuando la ingesta sobrepasa los 500 mg/da la absorcin disminuye porcentualmente. No obstante, las recomendaciones oficiales al respecto sealan que el contenido en colesterol de la dieta no debe nunca sobrepasar los 300 mg/da.
Acidos grasos saturados Como se ha visto anteriormente, los cidos grasos saturados carecen de dobles enlaces y les cuesta combinarse con otras molculas. Por este motivo, la mayor parte de las grasas saturadas se mantienen en estado slido a temperatura ambiente. Todas las grasas animales son altamente saturadas, excepto las del pescado y los mariscos, que son muy poliinsaturadas. Algunas grasas vegetales, como el aceite de coco y el de palma, son muy ricas en cidos grasos saturados. En numerosos estudios epidemiolgicos se ha comprobado que la ingesta de grasas saturadas aumenta los niveles de colesterol en sangre, especialmente los de la fraccin LDL. Aunque el mecanismo por el que este aumento se produce no est del todo esclarecido, parece ser que los cidos grasos saturados enriquecen los fosfolpidos de la membrana celular, interfiriendo con la funcin normal de los receptores LDL y reduciendo de esta forma la absorcin de las LDL por las clulas. Al reducirse la eliminacin de las LDL, su concentracin en la sangre es mayor. Los diferentes cidos grasos saturados tienen distintos comportamientos sobre los niveles de LDL-colesterol:
El cido Palmtico (C16:0) es el principal cido graso saturado presente en los alimentos de origen animal. Diferentes investigaciones han arrojado que incrementa los niveles de colesterol total y LDL, cuando sustituyen en la dieta a los hidratos de carbono u otro tipo de grasas.
El cido Mirstico (C14:0), aunque en menor medida que el palmtico, tambin aumenta la concentracin de colesterol total. La dieta mixta habitual contiene cantidades pequeas de cido mirstico, presente fundamentalmente en la mantequilla. E
El cido Esterico (C18:0) no eleva los niveles plasmticos de colesterol total, segn distintos estudios en animales y humanos, en contraste con otros cidos saturados. Este cido se metaboliza ms rpidamente hacia cido oleico que otras grasas saturadas.
La influencia del cido Larico (C12:0) sobre los niveles de colesterol en sangre todava no est clara, aunque se ha demostrado que el aceite de coco (rico en larico) aumenta ms los niveles de colesterol que la grasa de cordero. Los cidos grasos saturados de cadena corta (C10 y menor) apenas modifican la colesterolemia.
En la fotosntesis se distinguen dos fases, la luminosa y la de fijacin de CO2 Como cualquier proceso bioqumico la fotosntesis se puede representar por una ecuacin global, que en este caso resume una reaccin de xido-reduccin en la que el H2O cede electrones (en forma de hidrgeno) para la reduccin del CO2 a glcidos (CH2O)n, con liberacin de O2. CO2 + H2O luz O2 + (CH2O)n
Fase luminosa En esta etapa, tambin llamada fotodependiente, porque se da slo en presencia de luz, ocurren dos procesos bioqumicos necesarios para la sntesis de glucosa: la reduccin de NADP a NADPH.H con los hidrgenos de la molcula de agua y la sntesis de ATP. En la fase luminosa los pigmentos como las clorofilas a y b (figura 3), carotenos, xantofilas, ficoeritrinas y ficocianinas, que se encuentran asociados a la membrana tilacoidal, captan fotones y se excitan. Figura 3. Modelo de una molcula de clorofila. La regin inferior de la molcula corresponde a una cadena hidrofbica. En la parte superior se observa el anillo con dobles enlaces, y asociado a un Mg. Modificado de efn.uncor.edu Los fotones, o cuantos de luz, son cantidades discretas de energa que se propagan comoondas. La luz visible corresponde a la regin del espectro electromagntico comprendida entre 400 y 700 nm, regin en la que absorben los pigmentos presentes en las plantas y cianobacterias. Segn su longitud de onda (), los fotones tienen diferente cantidad de energa, de manera que cuanto menor es , mayor es la energa, que decrece desde la regin violeta a la roja (figura 4).
. Se representan las regiones del espectro y sus longitudes de onda. La longitud de onda est relacionada inversamente con la energa. Modificado de a-diba.net FOTOSNTESIS La diversidad de pigmentos permite absorber fotones en todo el rango del espectro visible Los pigmentos, con mximos de absorcin en longitudes de onda caractersticas (figura 5), se agrupan en la membrana tilacoidal en estructuras llamadas antenas, formadas por unas 200 300 molculas. Los distintos pigmentos absorben fotones con diferentes longitudes de onda, de manera que su diversidad le da a la planta la posibilidad de aprovechar todo el espectro visible (figura 5). El conjunto de pigmentos absorbe fotones en todo el rango visible del espectro. Estos pigmentos forman las antenas, mientras que el centro de reaccin est formado slo por clorofila. Tomado de monografas.comLos pigmentos de la antena se excitan con la energa de los fotones Los pigmentos contienen electrones fcilmente excitables. Los electrones ms externos pueden absorber la energa de un fotn y pasar a un estado excitado de mayor energa, y por lo tanto ms inestable. Un electrn perteneciente a un orbital molecular puede ser excitado y pasar a otro orbital molecular de mayor energa, por el aporte externo de energa. En el ejemplo usado la energa proviene de un fotn (h.). El mismo electrn al volver al estado basal libera la energa absorbida ().FOTOSNTESIS De esta forma, la energa se transmite entre molculas adyacentes en la antena, hasta alcanzar el centro de reaccin (figura 7). Como la transmisin de la excitacin electrnica acompaa la pendiente de energa, esta pasa de un pigmento cuyo pico de mxima absorcin es menor que el del pigmento siguiente, es decir, con menor requerimiento de energa para excitarse. Representacin de la antena y centro de reaccin de un fotosistema. Un fotn excita un electrn de un pigmento que funciona como captador de energa, que se transfiere de una molcula de pigmento a otra, hasta llegar al centro de reaccin. Cuando se excita la molcula de clorofila del centro de reaccin se oxida: pierde un electrn. Modificado de recursos.cnice.mec.esm La transferencia de energa desde la antena al centro de reaccin se realiza con una prdida mnima, en 10 seg. En este proceso de transferencia es determinante el ordenamiento preciso en la membrana de los tilacoides de los componentes de la antena y del centro de reaccin. La clorofila del centro de reaccin se oxida: pierde un electrn Las antenas, centros de reaccin y transportadores de electrones (figura 8) conforman los fotosistemas: el fotosistema I (PSI) o P700 (cuyo centro de reaccin tiene el mximo de absorcin a 700 nm) y el fotosistema II (PSII) o P680 (cuyo centro de reaccin tiene un mximo de absorcin a 680 nm). Cuando un fotn excita a un pigmento antena del PSII, un electrn se eleva a un potencial ms alto, y cuando vuelve al nivel basal la energa es transferida a otro pigmento antena y as sucesivamente hasta que se excita el centro de reaccin P680, que pierde un electrn. El electrn que pierde el P680 es captado por la feofitina, una clorofila sin Mg. A partir de la feofitina una cadena de transporte electrnico lleva al electrn al centro de reaccin del PSI (figura 8). El PSII sin el electrn queda como un radical catinico (con carga +). El PSII requiere de un electrn para llenar el hueco electrnico que se gener. En esto es clave el papel del agua, que mediante la fotlisis se descompone en 2H+ yEstos ltimos llegan, de a uno, al P680 oxidado (figura 8), mientras el oxgeno se libera a la atmsfera como O2. Cuando un fotn excita a un pigmento antena del PSI, ocurre lo mismo que en la antena del PSII. Un electrn se eleva a un potencial ms alto y comienza una transferencia de excitacin entre pigmentos de la antena, de manera que la energa es transferida al centro de reaccin P700. Este se oxida, y el electrn perdido es captado por un aceptor primario diferente al del PSII (figura 8). FOTOSNTESIS El electrn cedido por el P700 pasa por una secuencia de transportadores y despus de una serie de reacciones redox, reduce al NADP a NADPH.H (figura 8). El NADPH.H aporta en la fase de fijacin de CO2 el poder reductor necesario para reducir el CO2 a glucosa (figura 10). Observar que ocurre un continuo flujo de electrones desde el agua hasta el NADPH.H, segn se observa en el diagrama Z representado en la figura 8. En este flujo no cclico de electrones ocurren reacciones redox, donde pierden gradualmente energa que es usada para generar un gradiente protnico y sintetizar ATP. Cuando no es necesario el poder reductor ocurre el flujo cclico de electrones. En este, los electrones del P700 llegan a la ferredoxina, y en lugar de derivar al NADP vuelven nuevamente al P680 (figura 8):el flujo cclico permite producir ATP sin reducir al NADP.La energa de los fotones genera un gradiente protnico que permite la produccin de ATP La energa liberada por los electrones entre el PSII y el PSI es utilizada para bombear protones contra gradiente electroqumico, a travs de una membrana impermeable a H+, desde el estroma hacia el interior de los tilacoides (figura 9). Este bombeo de protones desde el estroma produce alcalinizacin en este compartimento (pH 8) y acidificacin del espacio intratilacoidal (pH 5). El potencial electroqumico que se genera a ambos lados de la membrana tilacoidal hace que los H+pasen al estroma, a travs de los canales protnicos CFo La energa liberada es usada para la conversin de ADP + Pi en ATP, proceso conocido
como fotofosforilacin. hace que el ATP y el NADPH.H se sinteticen del lado del estroma, donde sern consumidos en la fijacin de CO2. En mitocondrias y cloroplastos el mecanismo de sntesis de ATP se explica a travs de la generacin de un gradiente protnico. Una diferencia entre ambos es en la orientacin del gradiente: en la mitocondria se establece desde el espacio intermembrana a la matriz (de afuera a adentro), con la En el mismo diagrama se puede observar tanto el flujo cclico de electrones como el no cclico. Los electrones son excitados por la luz y pasan por una cadena transportadora de electrones a favor de un gradiente de energa hasta llegar al PSI, donde se haba generado un hueco electrnico. Desde el centro de reaccin del PSI sale un electrn por vez, que reduce al NADP, previo pasaje por una cadena de transportadores. Modificado de ecogenesis.com.arFOTOSNTESIS ATPasa dirigida hacia la matriz, mientras que en los cloroplastos es desde el espacio intratilacoidal hacia el estroma (de adentro hacia afuera), con la ATPasa dirigida hacia el estroma (figura 9). La energa liberada por los electrones es usada para bombear protones desde el estroma al espacio tilacoidal. Esto crea un gradiente electroqumico cuya energa permite que los protones se muevan a favor del gradiente a travs del complejo de CFo, yFase de fijacin de CO2La fijacin de CO2 y su reduccin a molculas glucdicas (CH2O)n se da en el estroma del cloroplasto a travs dereacciones reductivas, fuertemente endergnicas: el poder reductor (NADPH.H) y la energa (ATP) son producidos en la fase luminosa. Segn el mecanismo de fijacin de CO2 se establecen tres modelos fotosintticos, el de las plantas C3, el de las C4 y el de las CAM (Crassulacean Acid Metabolism). Las ms abundantes son las plantas C3, es decir, las que slo presentan ciclo C3, porque tanto las plantas C4 como CAM presentan adems un ciclo C3. Ciclo C3 o ciclo de Calvin: permite la conversin del CO2 en glcidos La energa y poder reductor generados en las reacciones fotoqumicas se usan para la conversin del CO2 en glcidos en el Ciclo de Calvin (figura 10). Este ciclo ocurre en el estroma del cloroplasto, dondese encuentran las enzimas que catalizan la conversin de dixido de carbono (CO2) en glcidos (triosas y glucosa). El CO2 ingresa a travs de los estomas (figura 2) y se une al aceptor, la ribulosa 1,5 bifosfato, por accin de la enzima rubisco (ribulosa bifosfato carboxilasa oxigenasa). La rubisco constituye aproximadamente el 50% de las protenas del cloroplasto y es la protena ms abundante en la naturaleza. El primer producto estable despus de la carboxilacin es el cido 3 fosfoglicrico (3PGA), un compuesto de 3 carbonos, al que se debe la denominacin de ciclo C3. En un ciclo se fijan 6 moles deCO2 a 6 moles de ribulosa 1,5 bifosfato, y se forman 12 moles de 1,3PGA con un consumo de 12 moles de ATP (figura 10). Para llegar a molculas de glcidos es necesaria la reduccin del 1,3PGA a GA3P, lo que se logra con 12 moles de NADPH.H (figura 10): observar que la direccin de las reacciones son las mismas que las ocurridas en la glucognesis. El ciclo se cierra con la regeneracin del aceptor, ribulosa 1,5P a partir de 10 moles de GA3P y el consumo de 10 moles de ATP: 10 GA3P x 3C = 30C (6 ribulosa 1,5P x 5C = 30C). FOTOSNTESIS Por otro lado los 2 moles de GA3P restantes pueden sintetizar glucosa en el cloroplasto y acumular almidn, o pueden ser transportado al citosol y ser usado en la gluclisis, o sintetizar glucosa y sacarosa, entre otros compuestos carbonados. En perodos de oscuridad las clulas fotosintticas se comportan como hetertrofas, y las clulas de tallos no fotosintticos y races siempre se comportan como hetertrofas, de manera que dependen para su nutricin de molculas glucdicas como la sacarosa, que llegan desde los tejidos fotosintticos va floema. La rubisco puede carboxilar u oxigenar a la ribulosa 1,5 P La enzima rubisco est formada por 16 subunidades, 8 mayores codificadas en el ADN del cloroplasto, y 8 menores codificadas en el ADN nuclear. La rubisco tiene la propiedad de unir en el mismo centro cataltico al CO2 o al O2, es decir tiene dos sustratos, y cataliza la carboxilacin o la oxigenacin: de ah su nombre ribulosa 1,5 bifosfatocarboxilasa oxigenasa. Cuando la enzima acta como carboxilasa fija CO2 a la ribulosa 1,5P dando comienzo el ciclo C3: se asimila el carbono. Cuando la enzima acta como oxigenasa, y oxigena a la ribulosa 1,5P, comienza un proceso conocido como fotorrespiracin figura 11. Si bien en la atmsfera hay un 21% de O2 y un 0.03% de CO2, la kM de la rubisco por el CO2 es 12 M, mientras que la kM por el O2 es 250 M. Esto hace que la asimilacin del CO2 sea mayor a al incorporacin de O2 a la ribulosa 1,5P, es decir, a la fotorrespiracin. FOTOSNTESIS La rubisco puede actuar como carboxilasa dando origen a la fijacin de CO2, es decir a la asimilacin de C, o como oxigenasa dando origen a la fotorrespiracin. La funcin de la fotorrespiracin es ms evidente en plantas en condiciones de estrs La capacidad oxigenasa de la rubisco tiende a ser vista como un mecanismo intil para la planta, porque desde el punto de vista de la eficiencia, no se aprecia una funcin til para la planta: la fotorrespiracin no conduce a la fijacin sino a la prdida de CO2, con gasto de ATP.
En condiciones normales de presin parcial de O2 y CO2 en atmsfera, hasta el 50% del carbono fijado por la fotosntesis puede perderse por fotorrespiracin. As, este proceso reduce la eficiencia fotosinttica en algunas plantas. La importancia de este proceso es ms comprensible en plantas en condiciones de estrs. A partir de resultados observados en plantas de tabaco, en las que se logr disminuir la velocidad de la fotorrespiracin mediante ingeniera gentica, se encontr que las plantas con niveles normales de fotorrespiracin toleraron ms la alta intensidad lumnica, respecto a las plantas con fotorrespiracin disminuida. Ciclo C4: las plantas con esta estrategia minimizan la fotorrespiracin Plantas como Paspalum, maz y caa de azcar, entre otras, tienen una va de captacin de CO2conocida como ciclo C4 que minimiza el proceso de fotorrespiracin. Este ciclo ocurre en el citosol de las clulas del mesfilo, mientras que el ciclo C3, que ocurre seguidamente al ciclo C4, lo hace en los cloroplastos de las clulas de la vaina (figura 12). En las plantas C4 la PEP carboxilasa incorpora el CO2 a una molcula de fosfoenolpiruvado (PEP), de 3 carbonos, que se transforma en oxalacetato (OAA), de 4 carbonos (figura 13). El OAA se transforma en malato (o en aspartato), que pasa a la clula de la vaina donde hay cloroplastos con rubisco. En la clula de la vaina el malato es decarboxilado y rinde CO2 y una molcula de 3C Mientras la molcula de 3C vuelve a la clula del mesfilo y regenera el PEP, con gasto de ATP, el CO2 es usado por la rubisco para carboxilar a la ribulosa 1,5P y dar comienzo al ciclo C3 (figura En las clulas de la vaina la concentracin CO2 puede ser tres o cuatro veces mayor que su concentracin en la atmsfera. De esta forma se evita buena parte de la actividad de oxigenasa de la rubisco, y la fotorrespiracin no es evidente: las plantas C4 minimizan la fotorrespiracin. Esta estrategia hace a las plantas C4 ms eficientes que las plantas C3, si se relaciona fotn absorbido con CO2 fijado, ambas en su condicin ptima. En las plantas C4 la condicin ptima se caracteriza por requerimientos de alta cantidad de luz, y elevada humedad y temperatura. En las clulas del mesfilo el CO2 se une al PEP por accin de la enzima PEPcarboxilasa (PEPc). El producto de esa reaccin, de 4C, pasa a la clula de la vaina y abastece de CO2 al ciclo de Calvin (o ciclo C3) y permite la regeneracin de PEP a partir del cido piruvato(de 3C). Tomado de FOTOSNTESIS Plantas CAM Como vimos hay tres tipos de plantas segn el mecanismo fotosinttico que presenten: las C3, las C4 y las CAM (Crassulacean Acid Metabolism). Ahora que estn todos los mecanismos fotosintticos descritos es importante hacer nfasis que, mientras que las plantas C3 slo presentan el ciclo C3 para fijar CO2, las plantas C4 y las CAM presentan otro ciclo previo al C3, pero todas cuentan con un ciclo C3. Las crasulceas, cactceas, bromeliceas, y orqudeas son plantas con CAM. La mayora de las plantas son C3, de todas formas las plantas C4 y las CAM estn presentes en la mayora de los ecosistemas, si bien las plantas CAM (Figura 14) estn adaptadas a condiciones extremas de temperatura y sequa, como ocurre en zonas desrticas. En las plantas CAM ocurren dos carboxilaciones en la misma clula, en diferente momento Durante la noche, cuando los estomas estn abiertos porque la temperatura desciende y la prdida de agua por evapotranspiracin es mnima, el CO2 ingresa a la planta y por accin de la enzima PEP carboxilasa se carboxila el PEP (Figura 15). Durante el da, en presencia de ATP y NADPH.H generados en la fase luminosa, ocurre la fijacin de CO2 por la rubisco a travs del ciclo C3: el ciclo C3 es comn a todos los modelos fotosintticos. Las plantas CAM fijan el CO2 al PEP en la primera carboxilacin y sintetizan oxalacetato, que se transforma en malato durante la noche. El malato que se almacena en las vacuolas de las clulas del parnquima es responsable del descenso del pH en las hojas durante la noche. El pH aumenta durante el da a medida que se consume el malato. La descarboxilacin del malato suministra CO2 a la rubisco, que cataliza la segunda carboxilacin con la puesta en marcha del ciclo C3, que ocurre durante el da (Figura 15). El piruvato generado por decarboxilacin del malato cierra el ciclo. En las plantas CAM la fijacin de CO2 est separada temporalmente en el da y la noche, mientras que en las plantas C4 la separacin es espacial, es decir, en dos tipos de clulas, las del mesfilo y las de la vaina. Las plantas CAM en general son facultativas, es decir tienen metabolismo CAM en determinadas condiciones ambientales y metabolismo C3 en otras. Si bien las plantas con esta modalidad de fijacin de CO2 son caractersticas de regiones con climas extremos, su distribucin geogrfica es extensa, lo que probablemente logren por la flexibilidad metablica que tienen para cambiar entre la modalidad C3 y CAM. Es ms, es comn que las plantas CAM funcionen como C3 en determinados momentos del da. FOTOSNTESIS Los estomas abiertos durante la noche permiten el ingreso de CO2, que se acumula como grupo carboxilo del oxalacetato. Durante el da la actividad de la fase luminosa genera ATP y NADPH.H, que permiten la reduccin del CO2 aportado por el malato a travs del ciclo C3.
Como la PEP carboxilasa y el CO2 liberado de la decarboxilacin del malato estn en el mismo compartimiento, si la enzima no estuviera regulada se generara un ciclo ftil de carboxilacin - decarboxilacin. Por esto es necesario que la PEP carboxilasa est regulada de tal manera que se eviteque el CO2 liberado de la descarboxilacin del malato sea nuevamente captado por el PEP para volver a para formar malato. Durante el da se evita esta situacin porque la PEP carboxilasa est inhibida por malato, y durante la noche la PEP carboxilasa se activa, incluso en presencia de malato, por modificacin covalente de la enzima: se fosforlila un residuo serina.
También podría gustarte
- Est. Const. Estado y GobiernoDocumento8 páginasEst. Const. Estado y GobiernoMartitaVillalobosAún no hay calificaciones
- Est. Const. Unidad II Estado y GobiernoDocumento8 páginasEst. Const. Unidad II Estado y GobiernoMartitaVillalobosAún no hay calificaciones
- Enfermedad de FabryDocumento4 páginasEnfermedad de FabryMartitaVillalobosAún no hay calificaciones
- Comida JaponesaDocumento4 páginasComida JaponesaMartitaVillalobosAún no hay calificaciones
- Rinitis PDocumento2 páginasRinitis PMartitaVillalobosAún no hay calificaciones
- Oracion 27 de OctubreDocumento3 páginasOracion 27 de OctubreXimena VizcainoAún no hay calificaciones
- Tema 2. Formación de Yacimientos Minerales.Documento42 páginasTema 2. Formación de Yacimientos Minerales.Yulimar CoronadoAún no hay calificaciones
- Proceso de Elaboración de La Coca-ColaDocumento3 páginasProceso de Elaboración de La Coca-ColaWilliam SaenzAún no hay calificaciones
- Guías de Ejercicios de Costos y Estandarización de Recetas - 9Documento5 páginasGuías de Ejercicios de Costos y Estandarización de Recetas - 9Paola GonzálezAún no hay calificaciones
- Analisis Capitulo 5 de Psicólogia y MasasDocumento7 páginasAnalisis Capitulo 5 de Psicólogia y MasasMike T. FrancoAún no hay calificaciones
- He LadosDocumento10 páginasHe LadosZiliany Coaquira QuilcaAún no hay calificaciones
- Clase 2 HigieneDocumento16 páginasClase 2 HigieneDARWIN MANUEL ALCIVAR RODRIGUEZAún no hay calificaciones
- Capitulo 14 GuytonDocumento29 páginasCapitulo 14 GuytonMaridiose Ruiz Barrera100% (2)
- Edwin Arnaldo Matt A ValdiviesoDocumento69 páginasEdwin Arnaldo Matt A ValdiviesoSindy NavaAún no hay calificaciones
- Emery Elementos de Genetica Medica 5aac85f71723dd3e0ab0472cDocumento9 páginasEmery Elementos de Genetica Medica 5aac85f71723dd3e0ab0472cMARIA ALEJANDRA PALOMINO ARCEAún no hay calificaciones
- Perdiguero Enfermerayantropologa SobreelpluralismoasitencialDocumento29 páginasPerdiguero Enfermerayantropologa SobreelpluralismoasitencialAlisson MartinezAún no hay calificaciones
- Sansom, CJ - El Gallo NegroDocumento198 páginasSansom, CJ - El Gallo NegroMonse Fndz100% (1)
- HOMBRÍA AL MÁXIMO - Cap 5 PDFDocumento7 páginasHOMBRÍA AL MÁXIMO - Cap 5 PDFJean Luka Bobadilla águilaAún no hay calificaciones
- Geomorfologia y RelieveDocumento4 páginasGeomorfologia y RelieveESMERALDAAún no hay calificaciones
- Base de Datos ConvenidosDocumento17 páginasBase de Datos ConvenidosMarcela MarAún no hay calificaciones
- P1 Presentación Casa Escorcia 2022Documento24 páginasP1 Presentación Casa Escorcia 2022The TopAún no hay calificaciones
- Revista+2020 2021+ +edit +ort 713 723Documento11 páginasRevista+2020 2021+ +edit +ort 713 723Miguel FigueroaAún no hay calificaciones
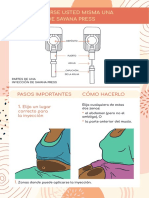
- Instrucciones Autoinyeccion Sayana PressDocumento5 páginasInstrucciones Autoinyeccion Sayana PressNeyda GuevaraAún no hay calificaciones
- Flujo Sobre Bancos de Tubos y Caída de PresiónDocumento2 páginasFlujo Sobre Bancos de Tubos y Caída de PresiónRAUL APAZA ROQUEAún no hay calificaciones
- Aps y Aps Renovada - Grupo 6.docx-CorrDocumento39 páginasAps y Aps Renovada - Grupo 6.docx-CorrLICHAAún no hay calificaciones
- Practica No.4 - RHJODocumento14 páginasPractica No.4 - RHJOOz HdzAún no hay calificaciones
- METANOLDocumento5 páginasMETANOLAnderson GalezzoAún no hay calificaciones
- Análisis BufferDocumento26 páginasAnálisis Bufferkarin santosAún no hay calificaciones
- Proyecto Final SalsarindoDocumento23 páginasProyecto Final SalsarindoAlejandrina Flores RodriguezAún no hay calificaciones
- Triptico QuemadurasDocumento2 páginasTriptico QuemadurasNORMA GONZALEZAún no hay calificaciones
- IDIFDocumento6 páginasIDIFCarolina Alexandra100% (2)
- El Tratamiento Endodóntico Desde La Óptica de La Odontología Restauradora 910-3516-1-PB PDFDocumento16 páginasEl Tratamiento Endodóntico Desde La Óptica de La Odontología Restauradora 910-3516-1-PB PDFgcanovas20072367Aún no hay calificaciones
- Aspectos Cuantitativo de La ElectrolisisDocumento13 páginasAspectos Cuantitativo de La ElectrolisisAngelis100% (1)
- Modelo Re Transistor Bipolar de JunturaDocumento2 páginasModelo Re Transistor Bipolar de JunturaLuis Abel Pilca SanungaAún no hay calificaciones
- HA210Documento1 páginaHA210JHOGAN JHONATHAN ALFONSO MENDOZAAún no hay calificaciones