También podría gustarte
- Clase AntioxidantesDocumento35 páginasClase AntioxidantesConstanza Belen GonzalezAún no hay calificaciones
- RADICALES LIBRES y NEUROTRANSMISORESDocumento10 páginasRADICALES LIBRES y NEUROTRANSMISORESFrancesca BarretoAún no hay calificaciones
- Resumen de Fisiologia Celular.Documento12 páginasResumen de Fisiologia Celular.Patricia De JesúsAún no hay calificaciones
- Citoplasma Celular 2008Documento36 páginasCitoplasma Celular 2008jhonny alexAún no hay calificaciones
- Mecanismo de Acción de AntisepticosDocumento2 páginasMecanismo de Acción de AntisepticosJose BetancourtAún no hay calificaciones
- Definicion de CitoesqueletoDocumento11 páginasDefinicion de CitoesqueletoAlberto VictorianoAún no hay calificaciones
- Ciclo Del Acido CitricoDocumento33 páginasCiclo Del Acido CitricoMabelizethAún no hay calificaciones
- Membrana CitoplasmaticaDocumento47 páginasMembrana CitoplasmaticaFernando MartinezAún no hay calificaciones
- Sistema de EndomembranasDocumento16 páginasSistema de EndomembranasAracelyIssaEnriquezTorrezAún no hay calificaciones
- CITOPLASMA InformeDocumento21 páginasCITOPLASMA InformeKatherin ArosteguiAún no hay calificaciones
- Mitocondria y CloroplastoDocumento10 páginasMitocondria y CloroplastoEduard Ramos VeraAún no hay calificaciones
- Envejecimiento CelularDocumento30 páginasEnvejecimiento CelularKaleb Alex MamaniAún no hay calificaciones
- MitocondriasDocumento26 páginasMitocondriasapi-3806371100% (1)
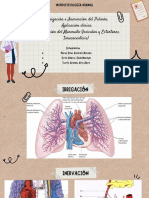
- Irrigación e Inervación Del Pulmón. Aplicación Clínica (Auscultación Del Murmullo Vesicular y Estertores, Toracocentesis) - Sem11Documento8 páginasIrrigación e Inervación Del Pulmón. Aplicación Clínica (Auscultación Del Murmullo Vesicular y Estertores, Toracocentesis) - Sem11Elida Marilyn Soto HilarioAún no hay calificaciones
- HemostasiaDocumento67 páginasHemostasiaMilis J NavasAún no hay calificaciones
- Aparato GolgiDocumento12 páginasAparato GolgiSoraida Sanchez Parra0% (1)
- 03 Mecanismos de Defensa Del Aparato Respiratorio Dr. Arnoldo Quezada L.Documento8 páginas03 Mecanismos de Defensa Del Aparato Respiratorio Dr. Arnoldo Quezada L.Miguel TapiaAún no hay calificaciones
- Funciones Del AmpcDocumento3 páginasFunciones Del AmpcRaulAún no hay calificaciones
- Anatomía y Fisiología CelularDocumento7 páginasAnatomía y Fisiología CelularDALVIC ALARCON50% (2)
- Retículo Endoplasmático RugosoDocumento4 páginasRetículo Endoplasmático RugosoC H Cruz CaballeroAún no hay calificaciones
- Transporte de O2 y Co2 Mecanismo de La Regulacion de La RespiracionDocumento13 páginasTransporte de O2 y Co2 Mecanismo de La Regulacion de La RespiracionLeslie Romina Paredes Cruz100% (1)
- Informe Sistema LinfaticoDocumento20 páginasInforme Sistema LinfaticomatiaslopezferrinAún no hay calificaciones
- Función de los orgánulos celularesDocumento24 páginasFunción de los orgánulos celularesAlbaniBriceñoAún no hay calificaciones
- Transporte CelularDocumento12 páginasTransporte CelularFelipeMendozaAún no hay calificaciones
- Guia VI - Metabolismo de Sustancias Nitrogenadas PDFDocumento11 páginasGuia VI - Metabolismo de Sustancias Nitrogenadas PDFnathalyAún no hay calificaciones
- FISIOPATOLOGÍA Sist DigestivoDocumento96 páginasFISIOPATOLOGÍA Sist DigestivoJUCARO2010Aún no hay calificaciones
- Lisosomas EnfermedadesDocumento4 páginasLisosomas EnfermedadesYasna Verdugo LeivaAún no hay calificaciones
- Organulos Membranosos y No MembranososDocumento19 páginasOrganulos Membranosos y No MembranososJesus Antonio Sanchez TzintzunAún no hay calificaciones
- Uniones Celulares Seminario 11Documento29 páginasUniones Celulares Seminario 11xiomara EusebioAún no hay calificaciones
- Grupo 11 Tejido Hematopoyético PreguntasDocumento2 páginasGrupo 11 Tejido Hematopoyético PreguntasMateoo06Aún no hay calificaciones
- Cuestionario de Glucidos y MetabolismoDocumento5 páginasCuestionario de Glucidos y MetabolismoIAGO SAMUEL DE SOUZA REISAún no hay calificaciones
- Cotransporte de Glucosa y Na - OliverDocumento14 páginasCotransporte de Glucosa y Na - OliverZavala Nuñez100% (1)
- Galactor Re ADocumento10 páginasGalactor Re AAnyhela Rodas GilAún no hay calificaciones
- Que Es La Nefrona y Como Es y Cual Es Su FuncionDocumento1 páginaQue Es La Nefrona y Como Es y Cual Es Su FuncionDiego Torres AltamiranoAún no hay calificaciones
- Reticulo Endoplasmatico Liso (Rel)Documento14 páginasReticulo Endoplasmatico Liso (Rel)Yeison PelufoAún no hay calificaciones
- Hormonas de Superficie CelularDocumento3 páginasHormonas de Superficie CelularAlisson Mejia Sanchez100% (1)
- Transporte Activo y PasivoDocumento9 páginasTransporte Activo y PasivoDavid Marin Antonio100% (2)
- Transporte de ProteinasDocumento9 páginasTransporte de ProteinasGise MathierAún no hay calificaciones
- CesarDocumento10 páginasCesarapi-3699557100% (1)
- Resumen Ciclo de KrebsDocumento2 páginasResumen Ciclo de KrebsJ Andres WolfAún no hay calificaciones
- Reticulo Endoplasmático LisoDocumento4 páginasReticulo Endoplasmático LisoDel Valle CarlitosAún no hay calificaciones
- Estructura y función de los ganglios linfáticosDocumento10 páginasEstructura y función de los ganglios linfáticosDiana Marcela AcostaAún no hay calificaciones
- MICROFILAMENTOSDocumento7 páginasMICROFILAMENTOSalmanza_1973Aún no hay calificaciones
- Diálisis en BioquímicaDocumento5 páginasDiálisis en BioquímicaGustavo VazquezAún no hay calificaciones
- EnzimasDocumento108 páginasEnzimasJohana GuzmánAún no hay calificaciones
- Fermentacion 3Documento10 páginasFermentacion 3rocioAún no hay calificaciones
- Tejido CartilaginosoDocumento36 páginasTejido CartilaginosoJessi Ramirez UAún no hay calificaciones
- Catalasa y OxidasaDocumento3 páginasCatalasa y OxidasaLiz De Mil ColoresAún no hay calificaciones
- GUIA BASICA DE HISTOLOGÍADocumento26 páginasGUIA BASICA DE HISTOLOGÍANath VelAún no hay calificaciones
- Fisiología RenalDocumento30 páginasFisiología RenalFatima AcostaAún no hay calificaciones
- Los Hidratos de CarbonoDocumento25 páginasLos Hidratos de CarbonoVivi Lorena Alvarado AriasAún no hay calificaciones
- Citoesqueleto UPAODocumento24 páginasCitoesqueleto UPAOCynthia GranadosAún no hay calificaciones
- Desarrollo de Vellosidades CorionicasDocumento11 páginasDesarrollo de Vellosidades CorionicasAlexia AlvaradoAún no hay calificaciones
- MitocondriaDocumento3 páginasMitocondriaValentina González IbarraAún no hay calificaciones
- Enzimas: Características y ClasificaciónDocumento27 páginasEnzimas: Características y ClasificaciónvictorAún no hay calificaciones
- DNP inhibe mitocondriasDocumento6 páginasDNP inhibe mitocondriasOskrAún no hay calificaciones
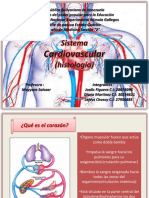
- Sistema Cardiovascular HistologiaDocumento26 páginasSistema Cardiovascular HistologiaAlbany ArteagaAún no hay calificaciones
- Lipidos MiaDocumento53 páginasLipidos MiaLuis Alberto Apaza ArceAún no hay calificaciones
- EosinofilosDocumento4 páginasEosinofilosEstefy JerezAún no hay calificaciones
- Vitaminas y Oxidorreductasas AntioxidantesDocumento30 páginasVitaminas y Oxidorreductasas AntioxidantesApril CollinsAún no hay calificaciones
- Artritis Septica LactanteDocumento5 páginasArtritis Septica LactanteShinvert Vera SanchezAún no hay calificaciones
- Neumonias en PediatriaDocumento47 páginasNeumonias en Pediatriapatonton120Aún no hay calificaciones
- Balance HidricoDocumento7 páginasBalance HidricoCarlos Estrada CampanaAún no hay calificaciones
- Numeros PosiblesDocumento100 páginasNumeros PosiblesShinvert Vera SanchezAún no hay calificaciones
- Centro de Salud ColomiDocumento34 páginasCentro de Salud ColomiShinvert Vera SanchezAún no hay calificaciones
- 5 - Embarazo AdolescenteDocumento10 páginas5 - Embarazo AdolescenteG3rm3nAún no hay calificaciones
- Exposicion Ulcera PepticaDocumento38 páginasExposicion Ulcera PepticaShinvert Vera SanchezAún no hay calificaciones
- PeruvianDocumento1 páginaPeruvianShinvert Vera SanchezAún no hay calificaciones
- Torneo PUBG PC 1000Bs CupoDocumento1 páginaTorneo PUBG PC 1000Bs CupoShinvert Vera SanchezAún no hay calificaciones
- Medicina TacticaDocumento95 páginasMedicina TacticaHugo Molas67% (3)
- Gotten Team Semana 04 SepDocumento1 páginaGotten Team Semana 04 SepShinvert Vera SanchezAún no hay calificaciones
- Refrigeracion PCDocumento1 páginaRefrigeracion PCShinvert Vera SanchezAún no hay calificaciones
- III Examen NeurologicoDocumento7 páginasIII Examen Neurologicohossito17Aún no hay calificaciones
- Ley Del Código de Etica de La Funcion Publica Ley27815Documento10 páginasLey Del Código de Etica de La Funcion Publica Ley27815Jc GuTiAún no hay calificaciones
- Esprue TropicalDocumento3 páginasEsprue TropicalShinvert Vera SanchezAún no hay calificaciones
- RadiologiaTrax MaDocumento53 páginasRadiologiaTrax MaShinvert Vera SanchezAún no hay calificaciones
- Guia Hemorragia Uterina AnormalDocumento12 páginasGuia Hemorragia Uterina Anormalhossito17Aún no hay calificaciones
- Ley Del Código de Etica de La Funcion Publica Ley27815Documento10 páginasLey Del Código de Etica de La Funcion Publica Ley27815Jc GuTiAún no hay calificaciones
- Cadena EpidemiologiaDocumento5 páginasCadena EpidemiologiaShinvert Vera SanchezAún no hay calificaciones
- Agitación PsicomotrizDocumento7 páginasAgitación PsicomotrizShinvert Vera SanchezAún no hay calificaciones
- Bioseguridad Tercer NivelDocumento11 páginasBioseguridad Tercer NivelShinvert Vera SanchezAún no hay calificaciones
- Planteamiento Del ProblemaDocumento3 páginasPlanteamiento Del ProblemaShinvert Vera SanchezAún no hay calificaciones
- 10 TrastornosalimenticiosDocumento35 páginas10 TrastornosalimenticiosShinvert Vera SanchezAún no hay calificaciones
- Cadena EpidemiologiaDocumento5 páginasCadena EpidemiologiaShinvert Vera SanchezAún no hay calificaciones
- HIPERPOTASEMIADocumento6 páginasHIPERPOTASEMIAShinvert Vera SanchezAún no hay calificaciones
- Un Paciente de 22 Años de Edad Del Chapare Acude A Consulta Medica Por Presentar FiebreDocumento3 páginasUn Paciente de 22 Años de Edad Del Chapare Acude A Consulta Medica Por Presentar FiebreShinvert Vera SanchezAún no hay calificaciones
- PREVENCIONDocumento5 páginasPREVENCIONShinvert Vera SanchezAún no hay calificaciones
- SexocraneoDocumento3 páginasSexocraneoShinvert Vera SanchezAún no hay calificaciones
- ObesidadDocumento11 páginasObesidadShinvert Vera SanchezAún no hay calificaciones
- Azul de TimolDocumento6 páginasAzul de TimolJUAN SEBASTIAN PIÑEROS GARCIA(PRACTICANTE)Aún no hay calificaciones
- Wiki-Polímeros: Análisis estructural de moléculas orgánicas y sus propiedadesDocumento12 páginasWiki-Polímeros: Análisis estructural de moléculas orgánicas y sus propiedadesKarla AguilarAún no hay calificaciones
- 2 Radiacion Del Cuerpo NegroDocumento5 páginas2 Radiacion Del Cuerpo NegroAnibal Lagos ParedesAún no hay calificaciones
- Nitrógeno LíquidoDocumento3 páginasNitrógeno LíquidoJuanCarlosFemeniasAún no hay calificaciones
- 9Documento31 páginas9nata2781Aún no hay calificaciones
- Instrumentación Industrial SEMANA 9Documento33 páginasInstrumentación Industrial SEMANA 9Marlen NarroAún no hay calificaciones
- Astronomía SOLUCIONES PDFDocumento7 páginasAstronomía SOLUCIONES PDFEvelyng EscalanteAún no hay calificaciones
- DMT producción poliéster resinasDocumento17 páginasDMT producción poliéster resinasjhanelaAún no hay calificaciones
- Factores Que Afectan La SolubilidadDocumento3 páginasFactores Que Afectan La SolubilidadCassandra GarciaAún no hay calificaciones
- 001ejer Mecanicafluidos PDFDocumento5 páginas001ejer Mecanicafluidos PDFJoha JácomeAún no hay calificaciones
- Acido Cítrico y GlucosaDocumento14 páginasAcido Cítrico y GlucosaDayanna Vasquez QuispeAún no hay calificaciones
- Cuarta Evaluación Cimentaciones 2021-0 Cordero Morales DalilaDocumento5 páginasCuarta Evaluación Cimentaciones 2021-0 Cordero Morales DalilaJean Pierre GarciaAún no hay calificaciones
- Laboratorio 1. Caida de PresionDocumento9 páginasLaboratorio 1. Caida de PresionKarol Castro SernaAún no hay calificaciones
- Topo Certamen 1BDocumento9 páginasTopo Certamen 1BAna María L. Soto LealAún no hay calificaciones
- Taller de Construccion Ambiental IDocumento10 páginasTaller de Construccion Ambiental IFujotakukillerAún no hay calificaciones
- Marco TeoricoDocumento2 páginasMarco TeoricoDiana Liz Gamarra ZeñaAún no hay calificaciones
- Taller de Movimiento Armonico SimpleDocumento7 páginasTaller de Movimiento Armonico SimpleLuis Alberto71% (7)
- Problemas T1Documento7 páginasProblemas T1Mauricio Arteaga SalinasAún no hay calificaciones
- Hidrocarburos Saturados e InsaturadosDocumento12 páginasHidrocarburos Saturados e InsaturadosGabriel GámezAún no hay calificaciones
- Control Optimo de La Inhibición y Estabilización de Las Lutitas y Arcillas en Los Fluidos de Perforacion Base AguaDocumento81 páginasControl Optimo de La Inhibición y Estabilización de Las Lutitas y Arcillas en Los Fluidos de Perforacion Base AguaMaritza salinas huanca100% (1)
- Tarea Extraclase Fisica TermodinamicaDocumento4 páginasTarea Extraclase Fisica TermodinamicaAlessa LexiAún no hay calificaciones
- Ficha 3 Periferia Atomica 1ra ParteDocumento11 páginasFicha 3 Periferia Atomica 1ra ParteMarcos Omo-Oxanguian MartinezAún no hay calificaciones
- Resultados Gelatina - Lab 5 - Tecno 3Documento7 páginasResultados Gelatina - Lab 5 - Tecno 3GFranco BlancasAún no hay calificaciones
- Monitoreo de Fuentes Presa HuacataDocumento7 páginasMonitoreo de Fuentes Presa Huacatawacho_189Aún no hay calificaciones
- Presiones hidrostáticas y empujes sobre objetos sumergidosDocumento2 páginasPresiones hidrostáticas y empujes sobre objetos sumergidosDeumek71% (7)
- Trabajo Practico N°3 - TP3 - Jorge Lorenzo Chuquimia CalderonDocumento12 páginasTrabajo Practico N°3 - TP3 - Jorge Lorenzo Chuquimia CalderonJORGE LORENZO CHUQUIMIA CALDERONAún no hay calificaciones
- Reporte de Fisica Ii #4 PDFDocumento21 páginasReporte de Fisica Ii #4 PDFVictoria AvilesAún no hay calificaciones
- Abp #2Documento16 páginasAbp #2Angie SierraAún no hay calificaciones
- Geodesía FísicaDocumento3 páginasGeodesía FísicaElias SeguielAún no hay calificaciones
- Comunicacion Oral y EscritaDocumento4 páginasComunicacion Oral y EscritaLilianaliz2008Aún no hay calificaciones