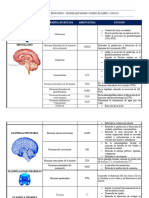
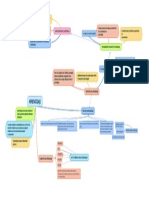
También podría gustarte
- Grijalva - Richard - Reproduccion y DesarrolloDocumento23 páginasGrijalva - Richard - Reproduccion y DesarrolloRichardAndresGrijalvaAún no hay calificaciones
- Catálogo de los copépodos (Crustacea: Copepoda: Calanoida y Cyclopoida) de cuerpos de agua temporales de Jalisco, México: Taxonomía y distribuciónDe EverandCatálogo de los copépodos (Crustacea: Copepoda: Calanoida y Cyclopoida) de cuerpos de agua temporales de Jalisco, México: Taxonomía y distribuciónAún no hay calificaciones
- Informe Practicas Jimmy Polo CorregidoDocumento31 páginasInforme Practicas Jimmy Polo CorregidoFlores JairAún no hay calificaciones
- Transgenicos en La AcuiculturaDocumento9 páginasTransgenicos en La AcuiculturaJen Krlos Hurtado BerrospiAún no hay calificaciones
- Ficha Técnica KampachiDocumento5 páginasFicha Técnica Kampachijesus lopezAún no hay calificaciones
- Reproduccion en PecesDocumento27 páginasReproduccion en PecesSaul GaleanoAún no hay calificaciones
- Camarón de MalasiaDocumento6 páginasCamarón de MalasiaROY EDSON ARPASI BARTOLOAún no hay calificaciones
- Cultivo de Boquichico AcuiculturaDocumento25 páginasCultivo de Boquichico AcuiculturaRosario Cristina Sosa YturriaAún no hay calificaciones
- Peces DietasDocumento10 páginasPeces DietasEdwards Milton Sullon Sullon100% (1)
- El MonengueDocumento26 páginasEl MonengueKarla Gurmendi Saavedra0% (1)
- Caracterizacion Cadena PisciculturaDocumento200 páginasCaracterizacion Cadena Pisciculturadiego100% (1)
- Estación Acuícola MoluscosDocumento8 páginasEstación Acuícola MoluscosSocorroSanchezFajardoAún no hay calificaciones
- 08 - Cultivo de Camarón de Agua Dulce PDFDocumento26 páginas08 - Cultivo de Camarón de Agua Dulce PDFEmiliano San100% (2)
- Ichthyophthiriasis en Peces Natvos Del Lago Titicaca en Puno (Peru)Documento6 páginasIchthyophthiriasis en Peces Natvos Del Lago Titicaca en Puno (Peru)Wilder Hernán Tisnado Angulo0% (1)
- Alimento para PecesDocumento2 páginasAlimento para PecesPamela CarbAún no hay calificaciones
- Alimento Vivo en La AcuaculturaDocumento7 páginasAlimento Vivo en La AcuaculturaJessamyn MedinaAún no hay calificaciones
- Cultivo de MicroalgasDocumento16 páginasCultivo de MicroalgasUliVanAún no hay calificaciones
- Disautonomia EquinaDocumento5 páginasDisautonomia EquinaEduFernándezMartínezAún no hay calificaciones
- TilapiaDocumento81 páginasTilapiaMike Wright100% (1)
- EER Ica Fernando KoechlinDocumento22 páginasEER Ica Fernando KoechlinMaría José Palape EncaladaAún no hay calificaciones
- TrichomonasDocumento5 páginasTrichomonaschicharito159Aún no hay calificaciones
- Manejo de EstanquesDocumento3 páginasManejo de EstanquesLuis-Miguel-VeizagaAún no hay calificaciones
- Rotiferos Levadura 2008Documento7 páginasRotiferos Levadura 2008ING. JULIO FERNANDO DIAZ MAMANIAún no hay calificaciones
- Concha de Abanico 1Documento40 páginasConcha de Abanico 1Miriam VelasquezAún no hay calificaciones
- Renibacterium SalmoninarumDocumento5 páginasRenibacterium SalmoninarumRommel Colque MoralesAún no hay calificaciones
- Mejoramiento en OvinosDocumento13 páginasMejoramiento en OvinosJason FernandezAún no hay calificaciones
- Cultivo del pulpo Octopus mimus enDocumento14 páginasCultivo del pulpo Octopus mimus enJunior Q. DavalosAún no hay calificaciones
- Requerimiento Nutricional en PecesDocumento16 páginasRequerimiento Nutricional en PecesCris Medina PoolAún no hay calificaciones
- Manual Basico de PatologiaDocumento52 páginasManual Basico de PatologiaKenji CoñesAún no hay calificaciones
- Disección y Marcaje de PecesDocumento10 páginasDisección y Marcaje de PecesJohnny Nuñez0% (1)
- Cultivo de Carpas y Otros Peces en EstanquesDocumento61 páginasCultivo de Carpas y Otros Peces en EstanquesAnonymous ZFbm2IyhOAún no hay calificaciones
- Glosario 04 de IctiopatologiaDocumento6 páginasGlosario 04 de IctiopatologiamichaelAún no hay calificaciones
- Truchas TriploidesDocumento8 páginasTruchas TriploidesAngel ParraAún no hay calificaciones
- Pob Laci OnesDocumento1 páginaPob Laci OnesAlex Valle SotoAún no hay calificaciones
- Biotecnología Aplicada A La AcuiculturaDocumento6 páginasBiotecnología Aplicada A La Acuiculturaingrid100% (1)
- Características de Los PoecílidosDocumento1 páginaCaracterísticas de Los PoecílidosJoiverDoriaPestanaAún no hay calificaciones
- Reproduccion y Desarrollo Embrionario en PecesDocumento25 páginasReproduccion y Desarrollo Embrionario en PecesLau Cq100% (1)
- Distomatosis HepaticaDocumento35 páginasDistomatosis HepaticaMaria Del Pilar GSAún no hay calificaciones
- Cultivo de Gamitana 2015 PDFDocumento54 páginasCultivo de Gamitana 2015 PDFFrancoGuerraGrandez100% (2)
- Acuicultura, Conceptos e HistoriaDocumento20 páginasAcuicultura, Conceptos e HistoriaemeadeaAún no hay calificaciones
- Protocolo de RotiferoDocumento19 páginasProtocolo de Rotiferoalexis diaz hurtadoAún no hay calificaciones
- Introducción A La Acuicultura - GUIADocumento8 páginasIntroducción A La Acuicultura - GUIAnervarialAún no hay calificaciones
- Masculinización de La "Tilapia Roja" FaltaDocumento30 páginasMasculinización de La "Tilapia Roja" FaltaDaner Quispe BalbinAún no hay calificaciones
- Clase ConsanguinidadDocumento16 páginasClase ConsanguinidadJeAn PieRcito OsiToAún no hay calificaciones
- Desarrollo Embrionario de Peces AnualesDocumento1 páginaDesarrollo Embrionario de Peces Anualestheblack08Aún no hay calificaciones
- Trabajo de Investigaciòn Semestral-Evaluaciòn de Recurso Hidrobiologico-Coco (Paralonchurus Peruanus) en El PerúDocumento25 páginasTrabajo de Investigaciòn Semestral-Evaluaciòn de Recurso Hidrobiologico-Coco (Paralonchurus Peruanus) en El PerúAlexander Sobrino Colán100% (1)
- Sistemas AcuicolasDocumento6 páginasSistemas AcuicolasSeirle Hammeir Calvo MercadoAún no hay calificaciones
- Epicin CatalogoDocumento6 páginasEpicin CatalogoEfrain Cayra100% (1)
- Aparato Reproductor Del CuyDocumento94 páginasAparato Reproductor Del CuyMayra Marcalla100% (1)
- Cadena de Valor en La Acuicultura PeruanaDocumento82 páginasCadena de Valor en La Acuicultura PeruanaJose Guillen Da CruzAún no hay calificaciones
- Cultivo Del Pangasius JuanjoseDocumento2 páginasCultivo Del Pangasius JuanjoseMarisol Cañate HerreraAún no hay calificaciones
- Alimentación de langostino con 12 dietas artificialesDocumento8 páginasAlimentación de langostino con 12 dietas artificialesDe Mendiburu MarisuAún no hay calificaciones
- 1337159678BienestarAnimal BajaDocumento171 páginas1337159678BienestarAnimal BajaamicrazyAún no hay calificaciones
- Taxonomia Del LangostinoDocumento60 páginasTaxonomia Del LangostinoArmando Martin Lira Rivas100% (1)
- Estrés Por Calor en Pollos de Engorde y Sus Efectos en La ProducciónDocumento25 páginasEstrés Por Calor en Pollos de Engorde y Sus Efectos en La ProducciónJorge Marino MedinaAún no hay calificaciones
- Cunicultura Como Alternativa de Produccion AnimalDocumento21 páginasCunicultura Como Alternativa de Produccion AnimalalfonsoenriquepinedaAún no hay calificaciones
- Actividad 2 - CyanoprokaryotaDocumento2 páginasActividad 2 - CyanoprokaryotaErick Diaz100% (1)
- Preparación de abono orgánico BocashiDocumento8 páginasPreparación de abono orgánico BocashiAB KHEYLA XIMENA RIVAS MARTINEZAún no hay calificaciones
- Leguminosas espontaneas de posible valor forrajero en ColombiaDe EverandLeguminosas espontaneas de posible valor forrajero en ColombiaAún no hay calificaciones
- Fotoperiodo en TruchasDocumento13 páginasFotoperiodo en TruchasJoselynCordovaPerezAún no hay calificaciones
- Sistema renina-angiotensina-aldosterona y su importancia fisiológicaDocumento5 páginasSistema renina-angiotensina-aldosterona y su importancia fisiológicaAleliz ZCAún no hay calificaciones
- Contextualizaci N de La Sacha InchiDocumento6 páginasContextualizaci N de La Sacha InchiAlejandro OviedoAún no hay calificaciones
- 700 - Rafael Marin PachecoDocumento129 páginas700 - Rafael Marin PachecoAlejandro OviedoAún no hay calificaciones
- Protocolo Alpha D3 en Dietas de Gallinas Con Diferentes Niveles de Calcio SuplementarioDocumento8 páginasProtocolo Alpha D3 en Dietas de Gallinas Con Diferentes Niveles de Calcio SuplementarioAlejandro OviedoAún no hay calificaciones
- Tarea Hidrocefalia CaninaDocumento2 páginasTarea Hidrocefalia CaninaAlejandro OviedoAún no hay calificaciones
- Bienestar de Las Aves de Corral en Los Países en DesarrolloDocumento15 páginasBienestar de Las Aves de Corral en Los Países en DesarrolloAlejandro Oviedo100% (1)
- Vigilancia CJDocumento50 páginasVigilancia CJAlejandro OviedoAún no hay calificaciones
- Laringotraqueitis Infecciosa AviarDocumento15 páginasLaringotraqueitis Infecciosa AviarAlejandro OviedoAún no hay calificaciones
- Cows PiracyDocumento2 páginasCows PiracyAlejandro OviedoAún no hay calificaciones
- Cows PiracyDocumento2 páginasCows PiracyAlejandro OviedoAún no hay calificaciones
- Links Inmuno y ParasitoDocumento2 páginasLinks Inmuno y ParasitoAlejandro OviedoAún no hay calificaciones
- Trabajo de Grado Yosidara Sepulveda 3 PDFDocumento51 páginasTrabajo de Grado Yosidara Sepulveda 3 PDFAlejandro OviedoAún no hay calificaciones
- Tarea Hidrocefalia CaninaDocumento2 páginasTarea Hidrocefalia CaninaAlejandro OviedoAún no hay calificaciones
- Práctica de Reflejos en AnimalesDocumento1 páginaPráctica de Reflejos en AnimalesAlejandro OviedoAún no hay calificaciones
- ÁCAROSDocumento35 páginasÁCAROSAlejandro OviedoAún no hay calificaciones
- Instrucciones TiDocumento1 páginaInstrucciones TiAlejandro OviedoAún no hay calificaciones
- Resumen Apuntes de EndocrinoDocumento5 páginasResumen Apuntes de EndocrinoAlejandro OviedoAún no hay calificaciones
- Práctica Toma de Temperatura AnimalDocumento2 páginasPráctica Toma de Temperatura AnimalMaria Fernanda Rojas LuqueAún no hay calificaciones
- Ascitis en Pollo de EngordaDocumento64 páginasAscitis en Pollo de EngordaAlejandro OviedoAún no hay calificaciones
- Reaccion y ReflejoDocumento5 páginasReaccion y ReflejoAlejandro OviedoAún no hay calificaciones
- Tarea Hidrocefalia CaninaDocumento2 páginasTarea Hidrocefalia CaninaAlejandro OviedoAún no hay calificaciones
- Reaccion y ReflejoDocumento5 páginasReaccion y ReflejoAlejandro OviedoAún no hay calificaciones
- ÁCAROSDocumento35 páginasÁCAROSAlejandro OviedoAún no hay calificaciones
- Las Leyes de NewtonDocumento4 páginasLas Leyes de NewtonAlejandro OviedoAún no hay calificaciones
- Funciones de las glándulas reproductivas masculinas en diferentes especiesDocumento35 páginasFunciones de las glándulas reproductivas masculinas en diferentes especiesAlejandro Oviedo100% (1)
- 01.FJLS 1de2 PDFDocumento131 páginas01.FJLS 1de2 PDFAlejandro OviedoAún no hay calificaciones
- Angiostrongylus vasorum infection in dogs: Presentation and risk factorsDocumento1 páginaAngiostrongylus vasorum infection in dogs: Presentation and risk factorsAlejandro OviedoAún no hay calificaciones
- Interacción de Los Factores Ambientales Con La Respuesta Del Comportamiento Productivo en Pollos de EngordeDocumento12 páginasInteracción de Los Factores Ambientales Con La Respuesta Del Comportamiento Productivo en Pollos de EngordemarioalfonsochoachiAún no hay calificaciones
- Introducción A R (T)Documento87 páginasIntroducción A R (T)Alejandro OviedoAún no hay calificaciones
- Taxonomia Animales DomesticosDocumento2 páginasTaxonomia Animales DomesticosAlejandro OviedoAún no hay calificaciones
- Proceso de Investigacion de AccidentesDocumento50 páginasProceso de Investigacion de Accidentesmartha_andrea_3Aún no hay calificaciones
- Anatomía cardíaca RMDocumento35 páginasAnatomía cardíaca RMCristian Goyeneche CasanovaAún no hay calificaciones
- Trabajo Colaborativo Uno Aporte IndividualDocumento4 páginasTrabajo Colaborativo Uno Aporte Individualluis barretoAún no hay calificaciones
- Cultivo de NaranjaDocumento28 páginasCultivo de NaranjaMilton100% (1)
- Primer Caretas, Cuento de Las 2000 PalabrasDocumento5 páginasPrimer Caretas, Cuento de Las 2000 PalabrasMercedes RiofrioAún no hay calificaciones
- PDF Informe de Laboratorio 1 de Mecanica de Suelos DLDocumento36 páginasPDF Informe de Laboratorio 1 de Mecanica de Suelos DLJesusAún no hay calificaciones
- APRENDIZAJEDocumento1 páginaAPRENDIZAJECristian ArrietaAún no hay calificaciones
- Planta de AmoniacoDocumento16 páginasPlanta de AmoniacoCesarIsacupeAún no hay calificaciones
- Gestión Administrativa de Recursos HumanosDocumento3 páginasGestión Administrativa de Recursos HumanosGenesis Neris LebronAún no hay calificaciones
- Carrefour CardsDocumento8 páginasCarrefour CardsLucía Martín75% (4)
- Mapa Conceptual Auditoria SaludDocumento1 páginaMapa Conceptual Auditoria SaludANDREA DIAZAún no hay calificaciones
- Triptico OxiurosDocumento3 páginasTriptico OxiurosLaura Perez LandaAún no hay calificaciones
- Fracturas de Pelvis. Presentacion.Documento41 páginasFracturas de Pelvis. Presentacion.Maria Fernanda Robles CelisAún no hay calificaciones
- Biotecnología NegraDocumento2 páginasBiotecnología NegraVALENTINA DAZA LONDOÑOAún no hay calificaciones
- Sistema Protección de Seguridad Industrial - TAREA2Documento16 páginasSistema Protección de Seguridad Industrial - TAREA2Mildred Cuello100% (5)
- HidrocortisonaDocumento4 páginasHidrocortisonaJennifer Marcatelli100% (1)
- Manejo Conductual en El Aula. XimenaDocumento27 páginasManejo Conductual en El Aula. XimenaXimena Covarrubias VarasAún no hay calificaciones
- Triptico ParasitologíaDocumento2 páginasTriptico ParasitologíaHéctor MillaAún no hay calificaciones
- Curso Contratos Nominas y Seguros SocialesDocumento322 páginasCurso Contratos Nominas y Seguros Socialesluckytauro100% (3)
- Neurofisiologia Del DolorDocumento21 páginasNeurofisiologia Del DolorCHARLES100% (1)
- Manual Grua TelescopicaDocumento10 páginasManual Grua Telescopicaap16484Aún no hay calificaciones
- HDS Chile Clorox® Disinfecting Mist Multi-Surface-Eucalyptus Pepermint-Lemongrass MandarinDocumento7 páginasHDS Chile Clorox® Disinfecting Mist Multi-Surface-Eucalyptus Pepermint-Lemongrass MandarinEvelyn OrellanaAún no hay calificaciones
- Monografía Cromo y CadmioDocumento18 páginasMonografía Cromo y CadmioKarina Irigoín SánchezAún no hay calificaciones
- Tipos de virus informáticos y sus característicasDocumento2 páginasTipos de virus informáticos y sus característicassarita torresAún no hay calificaciones
- Monografia 1 (5)Documento17 páginasMonografia 1 (5)Kristell PalominoAún no hay calificaciones
- 1 Ensayo Probeta en AceiteDocumento7 páginas1 Ensayo Probeta en AceiteElizabeth RendonAún no hay calificaciones
- Proyecto de TransformacionDocumento3 páginasProyecto de TransformacionBolsas BagsAún no hay calificaciones
- El paciente encamado: complicaciones y cuidados de enfermeríaDocumento24 páginasEl paciente encamado: complicaciones y cuidados de enfermeríaPaula del Río BlancoAún no hay calificaciones
- I Trim Cta 5ºDocumento45 páginasI Trim Cta 5ºRomel Rodrimo RenoguezAún no hay calificaciones
- Musculos de La LaringeDocumento21 páginasMusculos de La LaringeMaría GómezAún no hay calificaciones