Documentos de Académico
Documentos de Profesional
Documentos de Cultura
RPB v19n2 Machote v1
Cargado por
Leonardo RomeroDerechos de autor
Formatos disponibles
Compartir este documento
Compartir o incrustar documentos
¿Le pareció útil este documento?
¿Este contenido es inapropiado?
Denunciar este documentoCopyright:
Formatos disponibles
RPB v19n2 Machote v1
Cargado por
Leonardo RomeroCopyright:
Formatos disponibles
Rev. peru. biol.
ISSN 1561-0837
Universidad nacional Mayor de san Marcos
FacUltad de ciencias Biolgicas
volUMen 19 agosto, 2012 nMero 2
LIMA, PER
revista
PerUana de
Biologa
2
Rector
Dr. Pedro Atilio Cotillo Zegarra
Vicerrector de Investigacin
Dr. Bernardino Ramrez Bautista
Consejo Superior de Investigacin
Dr. Manuel Gongora Prado
Decana de la Facultad de Ciencias Biolgicas
Mag. Martha Valdivia Cuya
Directora Instituto de Investigacin en Ciencias Biolgicas Antonio Raimondi
Mag. Ins Miriam Grate Camacho
La Revista Peruana de Biologa es una publicacin cientfca
arbitrada, producida por el Instituto de Ciencias Biolgicas Antonio
Raimondi, Facultad de Ciencias Biolgicas de la Universidad Nacional
Mayor de San Marcos, Lima, Per, y auspiciada por el Vicerrectorado
de Investigacin. La Revista es publicada tres veces al ao (abril,
agosto y diciembre) y esta dedicada a la publicacin de artculos
cientfcos originales e inditos de las reas de Biodiversidad, Biotec-
nologa, Manejo ambiental, Ecologa y Biomedicina. La Revista publica
los trabajos realizados por acadmicos e investigadores nacionales y
extranjeros, en idioma espaol o ingls. Los trabajos recepcionados
son evaluados por rbitros segn criterios internacionales de calidad,
creatividad, originalidad y contribucin al conocimiento. La Revista
es publicada simultneamente en la pgina web de la Universidad.
Revista Peruana de Biologa -
Rev. peru. biol. - ISSN 1561-0837
Rev. peru. biol. - ISSN 1727-9933 (on line)
http://revistasinvestigacion.unmsm.edu.pe/index.php/rpb/
http://sisbib.unmsm.edu.pe/BVRevistas/biologia/biologiaNEW.htm
http://redalyc.uaemex.mx/
Copyright 2012
Facultad de Ciencias Biolgicas, UNMSM
Hecho el Depsito Legal 98-3017
Informacin adicional a:
Revista Peruana de Biologa
Facultad de Ciencias Biolgicas UNMSM
Ciudad Universitaria, Av. Venezuela Cdra. 34 s/n. Lima
Casilla Postal: 11-0058 Lima-11, Per.
Telfono 619-7000-1502 / Telefax 619-7000-1509
Editor Jefe, email: editor.revperubiol@gmail.com
revista PerUana de Biologa
Publicacin cientfca de la Facultad de Ciencias Biolgicas de la
Universidad Nacional Mayor de San Marcos
Resumida/Indizada (Abstracted/Indexed) en:
Peridica (ndice de Revistas Latinoamericanas en Ciencias),
LIPECS (Literatura Peruana en Ciencias de la Salud), Zoological
Record (BIOSIS), Scielo (Scientifc Electronic Library Online),
Index to American Botanical Literature (Te New York Botanical
Garden), BIOSIS Previews, Biological Abstracts (BIOSIS), ProQuest
(Biological Science Journals), Redalyc.
Foto en cartula: Nasa urens, Lomas de Villa Mara. Huber Trinidad
Editor jefe
Leonardo Romero
Comit Editor
Rina Ramrez
Carlos Pea
Csar Arana
Carlos Paredes
Comit consultivo en los recientes nmeros
Maximilian Weigend
Freie Universitt Berlin- Alemania
Sebastin Barrionuevo
Fundacin Miguel Lillo- Argentina
Luis Aguirre
Universidad Mayor de San Simn, Bolivia
Rodney Ramiro Cavichioli
Universidade Federal do Paran- Brasil
Hlio Ricardo da Silva
Universidade Federal Rural do Rio de Janeiro, Brazil
Carlos Frederico Duarte da Rocha
Universidade do Estado do Rio de Janeiro- Brasil
Fabrcio Rodrigues dos Santos
Universidade Federal de Minas Gerais- Brasil
Suzete Rodrigues Gomes
Instituto Butantan- Brasil
Davor Vrcibradic
Universidade do Estado do Rio de Janeiro- Brasil
Stefan Dennenmoser
University of Calgary, Canada
Roberto Melndez
Museo Nacional de Historia Natural- Chile
Berta Calonge Camargo
Pontifcia Universidad Javeriana, Bogot, Colombia
Sergio Solari
Universidad de Antioquia- Colombia
Jos Mara Gutirrez
Universidad de Costa Rica, Costa Rica
Finn Borchsenius
Aarhus University- Denmark
Julissa Roncal
Aarhus University- Denmark
Juan Rigoberto Tejedo Huaman
Universidad Pablo de Olavide- Espaa
Arnaud Bertrand
IRD. Institut de recherche pour le dveloppement- Francia
Francis Kahn
IRD. Institut de recherche pour le dveloppement, - Francia
Jean-Christophe Pintaud
Institut de Recherche pour le Dveloppement- Francia
Mutsunori Tokeshi
Kyushu University - Japon
Francisco Alonso Sols Marn
Universidad Nacional Autnoma de Mxico- Mxico
Mnica Romo
Asociacin Peruana para la Conservacin de la Naturaleza- Per
Renato Guevara-Carrasco
Instituto del Mar del Per- Per
Reynaldo Linares-Palomino
Universidad Nacional Agraria La Molina- Per
Marcel Gutirrez-Correa
Universidad Nacional Agraria La Molina - Per
Gretty K. Villena
Universidad Nacional Agraria La Molina - Per
Gerardo Lamas
Universidad Nacional Mayor de San Marcos- Per
Pablo Ramrez
Universidad Nacional Mayor de San Marcos- Per
Diana Silva
Universidad Nacional Mayor de San Marcos- Per
Juan Tarazona
Universidad Nacional Mayor de San Marcos- Per
Armando Yarlequ
Universidad Nacional Mayor de San Marcos- Per
Manuel Tantalen
Universidad Peruana Cayetano Heredia- Per
Nigel Pitman
Duke University- USA
Lucia Luna
University of Michigan- USA
Maria del Carmen Ulloa Ulloa
University of Missouri- USA
Blanca Len
University of Texas at Austin - USA
Kenneth Young
University of Texas at Austin USA
Paul Velazco
American Museum of Natural History, USA
125
(Contina...)
Revista PeRuana de Biologa
Volumen 19 Agosto, 2012 Nmero 2
Rev. peru. biol. ISSN 1561-0837
Contenido
HOMENAJE
127 A la memoria del doctor Abundio Sagstegui Alva (1932 2012)
In memoriam of Dr. Abundio Sagstegui Alva (1932 2012)
Eric F. Rodrguez Rodrguez y Blanca Len
Editorial
135 La Revista basurero
Te Journal landfll
Leonardo Romero
Trabajos originales
139 Una nueva especie de Salvia (Lamiaceae) del Norte de Per
A new species of Salvia (Lamiaceae) from Northern Peru
Abundio Sagstegui Alva y Eric F. Rodrguez Rodrguez
143 Una nueva especie de Deprea Raf. (Solanaceae) del Per
A new species of Deprea Raf. (Solanaceae) from Peru
Marco Antonio Cueva
e talo Francisco Trevio
149 Flora vascular de las lomas de Villa Mara y Amancaes, Lima, Per
Vascular fora from Villa Mara and Amancaes Lomas, Lima, Peru
Huber Trinidad, Elluz Huamn-Melo, Amalia Delgado y Asuncin Cano
159 Dieta de murcilagos flostmidos del valle de Kosipata, San Pedro, Cusco - Per
Diet of phyllostomid bats of Kosipata Valley, San Pedro, Cusco - Peru
Rossana Maguia, Jessica Amanzo y Luis Huamn
167 Reproduccin y alimentacin de Phytotoma raimondii, cortarrama peruana en El Gramadal, Ancash
Reproduction and food habits and of Phytotoma raimondii, Peruvian Plantcutter in El Gramadal, Ancash
Mario Rosina y Mnica Romo
175 Estructura de tallas de tortuga pico de loro Lepidochelys olivacea (Testudines: Cheloniidae) en Tumbes, Per
Size structure of Olive Ridley turtle Lepidochelys olivacea (Testudines: Cheloniidae) in Tumbes, Peru
Manuel Vera y Carlos A. Rosales
181 Patgenos introducidos al Per en post larvas de Litopenaeus vannamei importadas
Pathogen introduced to Peru by trade in postlarvae of Litopenaeus vannamei
Mervin Guevara y Rubn Alfaro
187 Optimizacin de la biorremediacin en relaves de cianuracin adicionando nutrientes y microorganismos
Optimization of cyanidation tailings bioremediation adding nutrients and microorganisms
Jasmin Hurtado y Arturo Berastain
193 Dosis nica de ciclofosfamida disminuye la calidad espermtica y el epitelio germinal masculino en ratones
Unique cyclophosphamide doses in male mice decrease the spermatic quality and germinal epithelium
Lyonal Acosta, Vctor Nez, Jonathan Vsquez, Jos Pino y Betty Shiga
199 Chemical composition of Apodanthera bifora, a Cucurbit of the dry forest in northwestern Peru
Composicin qumica de Apodanthera bifora, una cucurbitcea del bosque seco del noroeste peruano
Daniel Clark, Maribel Tupa, Andrea Bazn, Lily Chang and Wilfredo L. Gonzles
205 Diversidad gentica de papa nativa (Solanum spp.) conservada in situ en cinco regiones de Per
Genetic diversity of in situ conservation landraces potato (Solanum spp.) from fve regions of Peru
Julin Soto, Tulio Medina, Yeny Aquino y Rolando Estrada
126
Notas cientfcas
213 Descripcin del nido de Scolomys melanops (Rodentia, Cricetidae) y su relacin con Lepidocaryum tenue (Arecales, Arecaceae)
Description of Scolomys melanops nest (Rodentia, Cricetidae) and its relationship with Lepidocaryum tenue (Arecales, Arecaceae)
Edgardo M. Rengifo y Rolando Aquino
217 Ampliacin de la distribucin de Sephanoides sephaniodes (Trochilidae: Aves) en Argentina
Expansion of the distribution of Sephanoides sephaniodes (Trochilidae: Aves) in Argentina
Nancy Vernica Marinero, Ricardo Omar Cortez, Eduardo Alfredo Sanabria, Lorena Beatriz Quiroga
219 Herbarium Areqvipense (HUSA): informatizacin y representatividad de su coleccin
Herbarium Areqvipense (HUSA): computerization and representativeness of its collection
Italo F. Trevio, Diego A. Sotomayor, Marco A. Cueva, Rafael Perez, Laura Cceres, Daniel Ramos, Edgardo M. Ortiz y Victor Quipuscoa
Comentario
223 Una experiencia de gestin participativa de la biodiversidad con comunidades amaznicas
An experience on participatory management of biodiversity with Amazonian communities
Jos lvarez y Noam Shany
127
A la memoria del doctor Abundio Sagstegui Alva
Rev. peru. biol. 19(2): 127 - 134 (August 2012)
Rev. peru. biol. 19(2): 127 - 134 (Agosto 2012)
Facultad de Ciencias Biolgicas UNMSM
ISSN 1561-0837
A la memoria del doctor Abundio Sagstegui Alva (1932 2012)
Eric F. Rodrguez Rodrguez
1
y Blanca Len
2
In memoriam of Dr. Abundio Sagstegui Alva (1932 2012)
1 Herbarium Truxillense (HUT), Universidad Nacional de Trujillo. Jr. San Martn 392. Trujillo, Per. Email:efrr@unitru.edu.pe
2 Museo de Historia Natural UNMSM, Lima, Per. Plant Resources Center & Department of Geography and the Environment, University of
Texas at Austin, EE.UU. Email: leon@austin.utexas.edu
HOMENAJE
El 26 de mayo del 2012 tras un accidente automovilstico,
falleci en la ciudad de Trujillo, provincia Trujillo, departamento
La Libertad, Per, el profesor doctor Abundio Sagstegui Alva,
uno de los ms connotados cientfcos peruanos. Esta inesperada
desaparicin caus profundo pesar en la comunidad cientfca
nacional e internacional, pues nos haba dejado un verdadero
maestro, cientfco, e investigador botnico, adems de un ser
humano de gran calidad.
Abundio Sagstegui Alva naci en Guzmango (provincia
Contumaz, departamento Cajamarca) un 23 de octubre de
1932, en un hogar humilde y con precariedad econmica. Tras
vencer a la adversidad curs exitosamente sus estudios primarios
en el Centro Educativo N 108 de su tierra natal, obteniendo
una beca por ser el primer puesto en su promocin; hecho
que sirvi para realizar sus estudios secundarios en el Colegio
Nacional San Ramn de Cajamarca. Debido a su dedicacin y
ahnco al estudio obtuvo el premio de excelencia, continuando
su formacin acadmica en la Universidad Nacional de Trujillo
(UNT), donde se gradu como Profesor de Educacin Secun-
daria (1953), Bachiller en Ciencias Biolgicas (1958), ttulo
de Bilogo (1969) y Doctor en Ciencias Biolgicas en 1976.
En 1954 inici su carrera docente en el Centro Escolar de
Varones N 251 de Otuzco, La Libertad. Su carrera adminis-
trativa, docente y de investigacin en la Universidad Nacional
de Trujillo, la inici en la Facultad de Ciencias Biolgicas
como organizador del Herbario de la UNT (1956-1964), jefe
de prcticas en los cursos de Fitogeografa General y del Per y
de Ecologa Vegetal (1957-1962), jefe del Museo de Botnica
(1960-1988), profesor auxiliar tiempo completo (1963-1969),
profesor asociado (1969-1974) y profesor principal dedicacin
exclusiva (1974-1988) en los cursos de Botnica Fanerogmica
y Fitogeografa. Terminada su labor docente en la UNT pas a
formar parte de la Universidad Privada Antenor Orrego (UPAO)
donde lleg a ser vicepresidente acadmico de la Comisin Or-
ganizadora de dicha universidad (1988-1995), docente principal
a dedicacin exclusiva (1991-2005), fundador y director tanto
del Museo de Historia Natural, herbario HAO as como de la
revista Arnaldoa (1995-2005). En el ao 2006 retorn a su alma
Abundio Sagstegui (de pie) con ngel L. Cabrera (izq.) y Arnaldo Lpez (der.) en el Herbarium Truxillense
(HUT) de la Universidad Nacional de Trujillo (Foto cortesa familia Lpez lvarez)
128
Homenaje
Rev. peru. biol. 19(2): 127 - 134 (Agosto 2012)
mater, la UNT, donde fue nombrado Profesor Emrito Vitalicio
ese mismo ao y Profesor Investigador Honorario en Ciencias
Biolgicas el 2008; ejerciendo su investigacin cientfca en el
herbario HUT hasta los ltimos das de su vida.
El doctor Sagstegui en sus inicios recibi orientacin, gua e
infuencia de los doctores Nicols Angulo Espino (1888-1969)
y Arnaldo Lpez Miranda (1922-2010) de la UNT y del doctor
ngel Lulio Cabrera del herbario de La Plata (Argentina), a
quienes tuvo gratitud eterna, pues siempre los mencionaba en
el quehacer diario y en sus intervenciones orales y conferencias
magistrales. Conjuntamente con Nicols Angulo Espino, funda-
dor del Herbarium Truxillense (HUT) en 1941, y Arnaldo Lpez
Miranda fueron los pioneros de la ciencia botnica en el norte
del Per y formadores de innumerables botnicos (Rodrguez
2010). El doctor Sagstegui fue el primer curador (Lanjouw
& Stafeu 1964) y luego director en varias oportunidades del
Herbarium Truxillense (HUT), el herbario de la Universidad
Nacional de Trujillo.
Abundio Sagstegui se caracteriz por ser docente dedicado,
por sus magnfcas clases de botnica fanerogmica y ftogeogra-
fa, en las cuales con una buena pedagoga y didctica, basado en
el ejemplo y la comparacin, con las experiencias obtenidas en el
campo enseaba temas que todos hoy recordamos. El trabajo de
campo form parte de su quehacer, vnculo e inspiracin a nuevas
generaciones de estudiantes. En esas salidas l brindaba minu-
ciosas explicaciones, desde la herborizacin bsica de las plantas
hasta las descripciones y discusiones taxonmicas, as como sobre
el anlisis de los caracteres taxonmicos y la eleccin perfecta
para elaborar una clave dicotmica. A menudo mencionaba
debemos trabajar tcnica y cientfcamente, lo cual demostr
con la elaboracin de sus libros textos: Fitogeografa General y
del Per (1969-1986, seis ediciones), Manual de malezas de la
costa nor-peruana, Plantas invasoras de los cultivos de arroz y Flora
invasora de los cultivos del Per; todos basados estrictamente en
sus investigaciones y experiencias de campo de nuestra fora.
Incansable estudioso e investigador de la fora peruana,
destacando en taxonoma, sistemtica, forstica, conservacin
y etnobotnica con nfasis en el norte del Per; podramos
califcar a Abundio Sagstegui como el eterno apasionado de
las plantas, que indicaba constantemente a sus alumnos las
plantas son los seres que determinan la vida en el mundo. Estos
esfuerzos quedan demostrados en su legado de especies nuevas
y en el estudio de foras y frulas especialmente andinas. Las
fuentes de fnanciamiento para sus investigaciones eran de su
propio peculio, de las instituciones donde laboraba o de becas
de investigacin del extranjero a las cuales postul.
Con el Estudio Ecolgico de la Flora de los Alrededores de
Trujillo (1957) inicia su importante produccin cientfca; la
cual supera las cien publicaciones (ver Tabla 1). En 1964, con
Arnaldo Lpez Miranda (HUT) propusieron Arnaldoa peruviana
Lpez & Sagst. como una especie nueva para el norte del Per
(Lpez & Sagstegui 1964; Rodrguez 2010), y que sera el
inicio de su fructfera labor en taxonoma y como un augurio
a su contribucin al conocimiento de las Asteraceae. Tras su
perfeccionamiento en el Museo de La Plata (Argentina) bajo el
asesoramiento de ngel Lulio Cabrera en 1968, continu sus
estudios reconociendo nuevas entidades en los gneros Coreopsis
y Verbesina. Vale mencionar aqu, antes de su inters por las
compuestas que hoy todos conocemos, se dedic a las Cyperaceae
asesorado por el eminente ciperlogo argentino doctor Manuel
Barros durante los aos 1959-1960 (e.g.: Ciperceas nuevas para
el Per, Ciperceas de la Provincia de Trujillo). As mismo, mostr
inters por los helechos entre 1964 y 1968 bajo las recomen-
daciones y determinaciones taxonmicas de los doctores Rolla
M. Tryon y Alice F. Tryon, ambos de la Universidad de Harvard
(e.g.: Notas sobre Helechos Nor Peruanos I, Notas sobre Helechos
Nor Peruanos II). Tras su consolidacin como sinenterlogo,
reconoce cuatro gneros nuevos para la ciencia (Caxamarca,
Jalcophila, Parachionolaena y Pseudoligandra) y 97 especies nuevas
para la ciencia (Tabla 2). La trayectoria cientfca de Abundio
Sagstegui est perennizada en 32 taxones (31 especies y una
variedad) dedicadas en su honor por botnicos nacionales y
extranjeros (ver Tabla 3).
En su largo trajinar por diversos ambientes desde lomas
costeras, bosques a jalcas logr recolectar alrededor de 18000
nmeros botnicos depositados principalmente en los herba-
rios: HAO+, HUT, F, LP, MO, NY, US, USM (acrnimos en
Holmgren et al. 1990). Fue un preocupado del conocimiento
de la fora y la conservacin de los bosques montanos relictos del
norte peruano (e.g.: Inventario preliminar de la fora del Bosque
Monteseco, Inventario preliminar de la fora del Bosque Cachil),
de las lomas costeras [e.g.: Fitoecologa del Cerro Campana (Pro-
vincia de Trujillo)] y del conocimiento de nuestra ftodiversidad
divulgado a travs de los textos: Flora y Vegetacin de Contumaz,
Diversidad Florstica de Contumaz, Diversidad Florstica del
Norte de Per (Tomo I), Diversidad Florstica del Norte de Per.
Bosques Montanos (Tomo II), y Nuestra Biodiversidad. Sinopsis de
las Asteraceae del Per; as como en la organizacin de eventos
cientfcos de alto nivel (e.g.: IV Congreso Nacional de Botnica
en 1988, Simposio: Biodiversidad y Desarrollo del Norte del Per
en 1993, Simposio: Estrategias para Bioconservacin en el Norte
del Per en 1995, y I Congreso Internacional Diversidad Biolgica
y Cultural Andina el 2004, todos en Trujillo, Per).
Con especial sentimiento se recuerda (ER) una de sus ltimas
salidas al campo, el ao 2003, a las jalcas del departamento de
La Libertad en el marco del proyecto Floristic Inventory of the
Jalca Formations of Northern Peru, donde el maestro a pesar de
su avanzada edad segua liderando, enseando con sapiencia,
caminando por los cerros, y haciendo trabajo de campo como
cuando joven. Difcil ser olvidar su gran capacidad de trabajo y
disciplina. Siempre recordaba y parafraseaba lo mencionado por
uno de sus maestros, el doctor Nicols Angulo Espino (HUT),
Como quisiera que el sol se estacionara para seguir trabajando.
Era un convencido que cuando se tena resultados importan-
tes y nuevos conocimientos haba que darlos a conocer inmedia-
tamente. Seguro de contar con medios para tal divulgacin cre
y dirigi dos importantes revistas el Boletn de la Sociedad Bot-
nica de La Libertad en la UNT (1969) y Arnaldoa en la UPAO
(1991); esta ltima, una revista indexada y nombrada en honor
a su maestro Arnaldo Lpez Miranda (HUT) (Rodrguez 2010).
Por su impecable trayectoria cientfca, actividad intelectual y
labor realizada en educacin recibi numerosos premios, distin-
ciones honorifcas e incorporaciones a prestigiosas instituciones
acadmicas y botnicas [e.g.: En el siglo pasado: Diploma y
medalla de oro en reconocimiento a la permanente labor de
investigacin del Concejo Provincial de Trujillo (1984); Palmas
Magisteriales en el Grado de Maestro por parte del Ministerio
de Educacin (1985); Investigador Asociado al Field Museum,
129
A la memoria del doctor Abundio Sagstegui Alva
Rev. peru. biol. 19(2): 127 - 134 (August 2012)
Chicago, EE.UU. (desde 1983); Investigador Asociado de
Ohio State University, Columbus, EE.UU.; Profesor Emrito
de la UNT (1995); Decano del Colegio de Bilogos del Per
(1983-1984 y 1997-1999); considerado el Cientfco del Mi-
lenio (2000). En el presente siglo: Distincin de Primer Grado
del Libertador Simn Bolvar otorgada por la UNT (2004);
Diploma y Medalla de Honor en el Grado de Ofcial, otorgada
por el Congreso Nacional de la Repblica (2005); Premio Nacio-
nal de CONCYTEC 2005 (2006); nombramiento de Profesor
Emrito Vitalicio por la UNT (2006); Incorporacin como
Acadmico de Nmero a la Academia Nacional de Ciencias
del Per (2007); Premiacin por el Ministerio del Ambiente
por contribuir al conocimiento de la Biodiversidad de Per
(2008)]; Doctor Honoris Causa y como Catedrtico Honorario
de numerosas universidades peruanas, entre otros. Perteneci a la
Sociedad Botnica de La Libertad (Fundador), Sociedad Peruana
de Botnica (SPB), Sociedad Argentina de Botnica, Asociacin
Latinoamericana de Botnica (ALB), American Society of Plant
Taxonomic (ASPT), Botanical Society of America (BSA), Comi-
sin Directiva Organizacin Flora Neotrpica (OFN).
El valioso legado del Dr. Abundio Sagstegui Alva como
investigador en la Scientia Amabilis es y continuar siendo
reconocido por todo aquel interesado en la fora andina. La
trascendencia de sus obras cientfcas y humanas queda vigente
y esperamos que su ejemplo sirva a las prximas generaciones
de practicantes de esta ciencia.
Literatura citada
Holmgren, P.; N. H. Holmgren & L. C. Barnett. 1990. Index Herba-
riorum. Part I, The Herbaria of the world, 8
th
edition. New
York Botanical Garden. New York, NY, U.S.A.
Lanjouw, J. & F.A. Stafeu. 1964. Index Herbariorum. The Herbaria
of the World. Pat 1. Regnum Vegetabile. 31:187-188.
Lpez, A. & A. Sagstegui. 1964. Una nueva especie del gnero
Arnaldoa. Rev. Fac. Ciencias Biolgicas Univ. Nac. Tru-
jillo 1(1): 12-16.
Rodrguez, E. 2010. Dr. Arnaldo Lpez Miranda (26 de junio de
1922 - 7 de mayo de 2010). Arnaldoa 17(1): 147-153.
1957. Estudio Ecolgico de la Flora de los Alrededores de Trujillo. En el Concurso Cientfco, organizado por la Direccin de Extensin
Universitaria. Premio pecuniario y diploma.
1959. Ciperceas nuevas para el Per. Biota 2(19): 267-269.
1960. Las Ciperceas de la Provincia de Trujillo. Rev. Univ., 3
a
. poca. 9(17-18): 153-185.
1961. Clave provisional para la determinacin de las Juncceas Liberteas. Rev. Universitaria, 3 poca. 10(19 y 20): 69-72.
1963. Algunos aspectos Fitogeogrfcos de la Provincia de Trujillo. Revista del Crculo de Estudios Histrico-Geogrfcos Ral Porras
Barrenechea N 1. Rev. Univ. de Trujillo, Per.
1964. Notas sobre Helechos Nor Peruanos I. Rev. Fac. Ciencias Biolgicas, 1(1): 1-11.
1964. Una nueva especie del gnero Arnaldoa. Rev. Fac. Ciencias Biolgicas Univ. Nac. Trujillo 1(1): 12-16. (con A. Lpez M.).
1968. Notas sobre Helechos Nor Peruanos II. Rev. Fac. Ciencias Biolgicas Univ. Nac. Trujillo 1(4): 31-39.
1969. Una nueva especie de Verbesina (Compositae) del Per. Bol. Soc. Bot. La Libertad 1(2): 55-57.
1969. Cuatro especies nuevas de compuestas peruanas. Boletn de la Sociedad Argentina de Botnica 11(4): 240-250.
1970. Tres compuestas austroamericanas nuevas o crticas. Bol. Soc. La Libertad 2(1-2): 63-75.
1971. Una nueva especie de Coreopsis (Compositae) del Per. (con I. Snchez V.). Bol. Soc. Arg. Bot. 13(4): 337-340.
1973. Manual de las Malezas de la Costa Nor Peruana. 1. Edic. Edit. Talleres Grfcos de la Univ. Nac. de Trujillo, Per. 480 p. (Libro).
1974. Plantas invasoras de los cultivos de arroz. 1ra. Edic. Talleres Grafcos de la Univ. Nac. Trujillo. 142 p. (Libro).
1975. Contribucin de la Universidad nacional de Trujillo al conocimiento de nuestra Flora. Bol. Soc. Bot. La Libertad 7(1-2): 35-45.
1975. Nuevos taxa de Verbesina (Compositae) del Per. Bol. Soc. Arg. Bot. 16(3): 261-266.
1975. Contribucin al conocimiento de la fora adventicia en el Per. Rebiol 4 (1-2):11- 19. (con E. Alvitez I. y J. Mostacero).
1976. Dos nuevas especies del genero Verbesina (Compositae) del Peru. Hickenia 1(9): 41-47.
1978. Plantas invasoras poco conocidas. Bol. Soc. Bot. La Libertad 10(1-2): 13-19. (con J. Mostacero & E. Alvtez).
1980. Una nueva especie del gnero Browallia (Solanaceae). Hickenia 1(39): 215-218. (con O. Dios).
1980. Compuestas andino-peruanas nuevas para la ciencia. I. Bol. Soc. Argent. Bot. 19(1-2): 61-68.
1981. Un nuevo taxn del gnero Coreopsis (Compositae). Darwiniana 23(1): 223-225. (con I. Snchez V.).
1982. Dos nuevas especies del gnero Coreopsis (Compositae). Hickenia 1(50): 263-268.
1982. Compuestas andino-peruanas nuevas para la ciencia. II. Bol. Soc. Argent. Bot. 20(3-4): 303-316.
1982. Una nueva especie de Senecio L. (Compositae: Senecioneae) de Per. Hickenia 1(58): 313-316. (con E. Zardini).
1982. Dos nuevas especies de compuestas peruanas. Bol. Soc. Bot. La Libertad, 14 (1-2), 1982.
1983. Sobre la presencia de Ullucus aborigineus Bcher. En el Per. Bol. Soc. Bot. La Libertad, 13(1-2): 31-33. (con M. Mostacero L.).
1985. Compuestas andino peruanas nuevas para la ciencia III. Phytologia 57(6): 415-420.
1985. Four new species of Asteraceae from Peru. Britonia 37(1): 6-13. (con M.O. Dillon).
1985. New species and combinations in Belloa (Inuleae-Asteraceae). Phytologia 58(6): 392-400. (con M.O. Dillon).
1986. Jalcophila, a new genus of andean Inuleae (Asteraceae). Brittonia, 38(2): 162-167. (con M. Dillon).
1986. A new species of Solanum sect. Petota (Solanaceae) from northern Peru. Brittonia 38(1): 9-12. (con M.D. Whalen & S. Knapp).
1986. A new species of Monactis (Asteraceae) from northern Peru. Phytologia 61(1): 5-8.
1986. New species and status changes in Andean Inuleae (Asteraceae). Phytologia 59(4): 227-233. (con M.O. Dillon).
1986. A new species of Achyrocline (Inuleae-Asteraceae) from Peru. Phytologia 60(2): 107-110. (con M.O. Dillon).
Tabla 1. Produccin cientfca del Dr. Abundio Sagstegui Alva. Compilado por Eric F. Rodrguez y Blanca Len.
(contina...)
130
Homenaje
Rev. peru. biol. 19(2): 127 - 134 (Agosto 2012)
1986. Fitogeografa General y del Per. Sexta Edicin. Edicin. Talleres Grfcos de la U.N.T., Trujillo., Per. (Libro).
1987. A new species of Gynoxys (Asteraceae: Senecioneae) from nortnern Peru. Brittonia 39(4): 432-435. (con C. Tellez A.)
1988. New species of Llerasia (Asteraceae: Astereae) from Peru. Brittonia 40(4): 363-367. (con M.O. Dillon).
1988. Fitoecologa del Cerro Campana (Provincia de Trujillo). Bol. Soc. Bot. La Libertad 14(1-2): 1-47. (con J. Mostacero & S. Lpez).
1988. Dos nuevas especies de compuestas peruanas. Bol. Soc. Bot. La Libertad 14(1-2): 49-53.
1988. A new species of Stillingia (Euphorbiaceae) from northern Peru. Ann. Missouri Bot. Gard. 75(4): 1666-1668. (con I. Sanchez V. & M. Huft).
1988. Additions to South American Senecioneae (Asteraceae). Brittonia 40(2): 221-228. (con M.O. Dillon).
1989. Novedades en el gnero Coreopsis (Asteraceae). Bol. Soc. Bot. La Libertad 15(1): 23-28. (con I. Snchez V.).
1989. Vegetacin y fora de Contumaz. Editorial Libertad E.I.R.L. Trujillo. 78 p. (Libro).
1989. A new species of Symplocos (Symplocaceae) from northwestern Peru. Brittonia 41(1): 32-34. (con M.O. Dillon).
1990. Oligandra Less. revisited and the need for a new genus, Pseudoligandra (Asteraceae: Inuleae). (con M.O. Dillon). Taxon 39: 126-129.
1990. A new species of Ranunculus (Ranunculaceae) from Peru. Brittonia 42(3): 182-184. (con T. Duncan).
1991. A new species of Jaltomata (Solanaceae) from northwestern Peru. (con S. Knapp & T. Mione). Brittonia 43(3): 181-184.
1991. Family Asteraceae: Part V. Fieldiana. Botany: New Series, N 26: 1- 70. (con M. O. Dillon).
1991. Compuestas andino-peruanas nuevas para la Ciencia IV. Arnaldoa 1(1): 1-10.
1991. Inventario preliminar de la fora del bosque de Monteseco. (con M.O. Dillon). Arnaldoa 1(1): 35-52.
1991. Una nueva especie de Chuquiraga (Asteraceae-Mutisieae) del Norte del Per. Arnaldoa 1(2): 1-4. (con I. Snchez V.)
1991. Sinopsis de los Gneros de Gnaphaliinae (Asteraceae-Inuleae) de Sudamrica. Arnaldoa 1(2): 5-91. (con M.O. Dillon).
1993. Una nueva especie de Trixis (Mutisieae: Asteraceae) del norte del Per. (con M.O. Dillon). Arnaldoa 1(3): 9-13.
1993. Revisin del gnero Arnaldoa (Compositae, Barnadesioideae), gnero endmico del Norte del Per. Arnaldoa 1(4): 9-21. (con T.F.
Stuessy).
1993. Variacion aloenzimtica en la rara especie endmica peruana Chuquiraga oblongifolia (Asteraceae). Arnaldoa 1(4): 73-76. (con D.
Crawford, T.F. Stuessy & I. Snchez V.)
1993. Flora Invasora de los cultivos del Per. CONCYTEC, Trujillo, Per. 539 p. (Libro).
1994. Flora endmica de los Andes Norperuanos. Arnaldoa 2 (1): 43-64.
1994. Estudios en la tribu Liabeae (Asteraceae) en Per: I. Revision de Ferreyranthus. Arnaldoa 2(2): 7-23. (con M.O. Dillon).
1994. Estudios en la tribu Liabeae (Asteraceae) en Per: II. Una nueva especie de Oligactis procedente del norte del Per y Sur del Ecuador.
Arnaldoa 2(2): 25-30. (con M.O. Dillon).
1994. Estudios en la tribu Liabeae (Asteraceae) en Per: III. Una nueva especie de Chrysactinium del norte del Per. Arnaldoa 2(2): 31-35.
(con M.O. Dillon).
1994. Compuestas andino-peruanas nuevas para la ciencia V. Arnaldoa 2(2): 37-45. (con I. Snchez & D. Crawford).
1995. Diversidad Florstica de Contumaz. Editorial Libertad E.I.R.L.. 203 p. (Libro).
1995. Floristic inventory and biogeographic analysis of montane forest in nortern Peru. Edited by S. Churchill et al. In Biodiversity and
Conservation of Neotropical Montane. 251-269. The New York Botanical Garden. (con M.O. Dillon, I. Snchez , S. LLatas, N. Hensold).
1995. Compuestas andino-peruanas nuevas para la ciencia. VI. Arnaldoa 3(2): 7-12. (con S. Leiva & P. Lezama).
1995. Inventario preliminar de la fora del Bosque de Cachil. Arnaldoa 3(2): 19-34. (con S. Leiva, P. Lezama, N. Hensold & M.O. Dillon).
1996. Compuestas andino-peruanas nuevas para la ciencia. VII. Arnaldoa 4(1): 7-13.
1996. El gashmin (=Eugenia quebradensis Mc Vaugh) un nuevo recurso alimenticio. Arnaldoa 4(1): 47-56.
1996. Plantas promisorias: La Taya o Tara. Arnaldoa 4(1): 57-65. (con P. Lezama & E. Marin).
1996. Revision of the dioecius genus Chersodoma Phil. (Senecioneae, Asteraceae), including a new species and status change. Brittonia 48
(4): 582-604. 1996 (con M.O. Dillon).
1997. A new species of Eucrosia (Amaryllidaceae) from northern Peru. Sida 17(4): 761-764. (con A.W. Meerow).
1998. Una nueva especie de Critoniopsis (Vernonieae: Asteraceae) de Cajamarca, Per. Arnaldoa 5(1): 19-24. (con M.O. Dillon).
1998. Una breve revisin del gnero Vernonia (sensu lato) del Per. Arnaldoa 5(1): 25-33. (con M.O. Dillon).
1998. Seis nuevas especies de Verbesina (Asteraceae, Heliantheae) de los Andes del Per. Arnaldoa 5(1): 35-50. (con V. Quipuscoa).
1998. Una nueva especie de Chrysophyllum (Sapotaceae) con frutos comestibles del noroeste del Per. Arnaldoa 5(1): 67-75. (con M.O. Dillon).
1998. Tres nuevas especies de Verbesina (Asteraceae: Heliatheae) de los Andes del Per. Arnaldoa 5(2): 141-149. (con V. Quipuscoa).
1999. Caxamarca, a new monotypic genus of Senecioneae (Asteraceae) from Northern Peru. Novon 9:156-161. (con M.O. Dillon).
1999. Diversidad forstica del norte del Per. Tomo I. Edit. Grafcart, Trujillo, Per. 228 p. (con. M.O. Dillon, I. Snchez, S. Leiva & P. Lezama).
1999. Verbesina ancashensis y V. crassicephala (Asteraceae: Heliantheae): dos nuevas especies del Per. Arnaldoa 6 (1): 41-48.
1999. La diversidad biolgica y El Nio 1997 98 en el norte del Per. El Nio 1997-98 y su Impacto en los Ecosistemas Marino y Te-
rrestre. J. Taramona & E. Castillo (Eds.). Rev. peru. biol. (Volumen Extraordinario): 183-186.
2001. Una nueva especie de Critoniopsis (Vernonieae: Asteraceae) del Norte de Per. Arnaldoa 8(1): 25 30. (con M. O. Dillon).
2001. Verbesina citrina (Heliantheae: Asteraceae), una nueva especie de Cajamarca, Per, Arnaldoa 8 (1): 31 36. (con M. Zapata).
2001. Tribal classifcation and diversity in the Asteraceae of Peru. Arnaldoa 8(2): 25 44. (con M.O.Dillon).
2001. Nuevo nombre para una especie de Chrysophyllum (Sapotaceae) del norte del Per. (con M.O. Dillon). Arnaldoa 8(2): 57-58.
2002. Biogeografa, diversidad forstica y endemismos en dos tipos de bosque en el norte de Per. Arnaldoa 9(2):111120. (con M.O. Dil-
lon, I. Snchez, G. Iberico, M. Zapata & M.L.Kawasaki).
2003. Diversidad forstica del norte del Per. Bosques Montanos. Tomo II. Edit. Grafcart, Trujillo, Per. 305 p. (con I. Snchez, M. Zapata
& M.O. Dillon).
2003. Coreopsis poloe (Asteraceae: Heliantheae), una nueva especie del Norte de Per. Arnaldoa 10(2): 15-18. (con M. Zapata).
2004. Coreopsis gracilis (Asteraceae: Heliantheae) una nueva especie del Norte de Per Arnaldoa 11(1): 25-28. (con I. Snchez).
2004. Una nueva especie de Puya (Bromeliaceae) del Norte de Per. Arnaldoa 11(2): 29 36. (con. M. Zapata & M.O.Dillon)
2007. Nuevo nombre para una especie de Coreopsis (Asteraceae: Heliantheaceae) del Norte del Per. Sciendo 10(1): 87. (con I. Snchez V.).
2007. Plantas Promisorias: El mito o Papaya Silvestre. Rev. Innova Norte 1(1):109-119. (con E. Rodrguez & S. Arroyo).
Tabla 1. Continuacin.
(contina...)
131
A la memoria del doctor Abundio Sagstegui Alva
Rev. peru. biol. 19(2): 127 - 134 (August 2012)
2007. Estudio Fitoqumico Preliminar en Tessaria intergrifolia. Ruiz & Pav. Ciencia y Salud 1 (1): 2-4. (con F. Prez & F. Rodrguez).
2007. Actividad antileishmaniasica in Vitro del extracto metanlico de las fores de Tessaria intergrifolia Ruiz & Pav. (Asteraceae). Pueblo
Continente 18 (1): 101-108. (con E. Marn & F. Prez).
2008. Una nueva especie de Ophryosporus (Eupatorieae: Asteraceae) para el Per. Rev. peru. biol. 15(1): 21-23 (con E. Rodrguez).
2008. General overview of Tillandsia subgenus Tillandsia in Peru: The three-pinnate species and the case of two endemic species. Rev. peru.
biol. 15(1): 25-30 (con B. Len).
2009. Nuestra Biodiversidad. Sinopsis de las Asteraceae del Per. 1ra Edicin. Ed. Grafcart, Trujillo, Per. 51 p. (Libro).
2009. Una nueva especie de Pentacalia (Senecioneae: Asteraceae) del Norte de Per. Rev. peru. biol. 16(2): 157-162. (con E. Rodrguez).
2009. Passifora tripartita var. mollisima (Passiforaceae) una especie promisoria de los Andes. Rev. Innova Norte. 2(1):11-29.(con E. Rodr-
guez & S. Arroyo).
2010. Una nueva especie de Axinaea (Melastomataceae: Merianieae) del Norte del Per. Rev. peru. biol. 17(2): 145-150. (con S. Arroyo &
E. Rodrguez).
2012. Una nueva especie de Salvia (Lamiaceae) del Norte de Per. Rev. peru. biol. 19(2): 139 - 142 (con E. Rodrguez).
Tabla 1. Continuacin.
Gneros
1. Caxamarca Sagst. & M.O. Dillon -PERU
Caxamarca sanchezii M.O. Dillon & Sagst.
2. Jalcophila M.O. Dillon & Sagst. ECUADOR, PERU
Jalcophila ecuadorensis M.O. Dillon & Sagst.
Jalcophila peruviana M.O. Dillon & Sagst.
3. Parachionolaena M.O. Dillon & Sagst -COLOMBIA
Parachionolaena colombiana (S.F. Blake) M.O. Dillon & Sagst.
4. Pseudoligandra M.O. Dillon & Sagst. BRASIL
Pseudoligandra chrysocoma (Wedd.) M.O. Dillon & Sagst.
Especies
1. Achyrocline peruviana M.O. Dillon & Sagst.
2. Arnaldoa peruviana Lpez & Sagst.
3. Axinaea wurdackii Sagst., S.J. Arroyo & E. Rodr.
4. Belloa plicatifolia Sagst. & M.O. Dillon
5. Belloa spathulifolia Sagst. & M.O. Dillon
6. Belloa turneri Sagst. & M.O Dillon
7. Browallia acutiloba Sagst. & Dios
8. Caxamarca sanchezii M.O. Dillon & Sagst.
9. Chersodoma deltoidea Sagst. & M.O. Dillon
10. Chrysactinium breviscapum Sagst. & M.O. Dillon
11. Chrysophyllum edule Sagst. & M.O. Dillon, nom. ileg. (=Chrysophyllum contumacense Sagst. & M.O. Dillon)
12. Chuquiraga oblongifolia Sagst. & Snchez Vega
13. Coreopsis breviligulata Sagst. & Snchez Vega
14. Coreopsis cajamarcana Sagst. & Snchez Vega
15. Coreopsis canescentifolia Sagst.
16. Coreopsis celendinensis Sagst. & Snchez Vega
17. Coreopsis dentifolia Snchez Vega, Sagst, & D.J. Crawford
Tabla 2. Gneros y especies descritas por el doctor Abundio Sagstegui Alva. Compilado por Eric F. Rodrguez y Blanca Len.
(contina...)
132
Homenaje
Rev. peru. biol. 19(2): 127 - 134 (Agosto 2012)
18. Coreopsis dilloniana Snchez Vega, Sagst. & D.J. Crawford
19. Coreopsis ferreyrae Sagst. & Snchez Vega
20. Coreopsis gracilis Sagst. & Snchez Vega, nom. ileg. (=Coreopsis mollicula Sagst & Snchez Vega)
21. Coreopsis helleborifolia Snchez Vega, Sagst. & Crawford
22. Coreopsis lopez-mirandae Sagst.
23. Coreopsis obovatifolia Sagst.
24. Coreopsis peruviana Sagst.
25. Coreopsis pervelutina Sagst.
26. Coreopsis poloe Sagst. & Zapata
27. Critoniopsis ayabacensis Sagst. & M.O. Dillon
28. Critoniopsis oblongifolia Sagst. & M.O. Dillon
29. Dasyphyllum brevispinum Sagst. & M.O. Dillon
30. Dasyphyllum cabrerae Sagst.
31. Eucrosia calendulina Meerow & Sagst.
32. Gamochaeta monticola M.O. Dillon & Sagst.
33. Gamochaeta oreophila M.O. Dillon & Sagst.
34. Gynoxys colanensis M.O. Dillon & Sagst.
35. Gynoxys congestifora Sagst. & M.O. Dillon
36. Gynoxys dilloniana Sagst. & C. Tllez
37. Gynoxys lopezii M.O. Dillon & Sagst.
38. Jalcophila ecuadorensis M.O. Dillon & Sagst. - ECUADOR
39. Jalcophila peruviana M.O. Dillon & Sagst.
40. Jaltomata whalenii S. Knapp, Mione & Sagst.
41. Llerasia pascoensis Sagst. & M.O. Dillon
42. Llerasia sanmartinensis Sagst. & M.O. Dillon
43. Loricaria ollgaardii M.O. Dillon & Sagst. - ECUADOR
44. Monactis rhombifolia Sagst. & M.O. Dillon
45. Oligactis cuatrecasasii M.O. Dillon & Sagst.
46. Onoseris chrysactinioides Sagst. & M.O. Dillon
47. Onoseris linearifolia Sagst.
48. Ophryosporus marchii Sagst. & E. Rodr.
49. Pentacalia barbourii M.O. Dillon & Sagst.
50. Pentacalia vallejiana Sagst, & E. Rodr.
51. Puya tyleriana Sagst., Zapata & M. O. Dillon
52. Ranunculus lambayequensis T. Duncan & Sagst.
53. Salvia vargas-llosae Sagast. & E. Rodr.
54. Senecio miniauritus Sagst. & M.O. Dillon
55. Senecio piurensis Sagst. & Zardini
56. Solanum guzmanguense Whalen & Sagst.
57. Stillingia parvifolia Snchez Vega, Sagst. & Huft
58. Symplocos incahuasensis Sagst. & M.O. Dillon
59. Trixis montesecoensis Sagst. & M.O. Dillon
60. Verbesina acosta-solisii Sagst. ECUADOR
61. Verbesina albissima Sagst.
62. Verbesina amplitifolia Sagst. & Quipuscoa
63. Verbesina ancashensis Sagst. & Quipuscoa
64. Verbesina andina Sagst.
65. Verbesina ayabacensis Sagst.
(contina...)
Tabla 2. Continuacin.
Especies
133
A la memoria del doctor Abundio Sagstegui Alva
Rev. peru. biol. 19(2): 127 - 134 (August 2012)
66. Verbesina aypatensis Sagst & Quipuscoa
67. Verbesina brevilingua Sagst.
68. Verbesina brunnea Sagst & Quipuscoa
69. Verbesina cabrerae Sagst.
70. Verbesina cajamarcensis Sagst.
71. Verbesina capituliparva Sagst.
72. Verbesina chachapoyensis Sagst. & Quipuscoa
73. Verbesina citrina Sagst. & Zapata
74. Verbesina contumacensis Sagst.
75. Verbesina crassicephala Sagst. & Quipuscoa
76. Verbesina dilloniana Sagst.
77. Verbesina ecuatoriana Sagst. - ECUADOR
78. Verbesina fuscicaulis Sagst.
79. Verbesina hexantha Sagst.
80. Verbesina huancabambae Sagst. & Quipuscoa
81. Verbesina huaranchaliana Sagst.
82. Verbesina leivae Sagst. & Quipuscoa
83. Verbesina lopez-mirandae Sagst.
84. Verbesina monactioides Sagst., S. Leiva & Lezama
85. Verbesina ochroleucotricha Sagst.
86. Verbesina otuzcensis Sagst. & Quipuscoa
87. Verbesina paucicephala Sagst.
88. Verbesina pauciramea Sagst., S. Leiva & Lezama
89. Verbesina pentalobifolia Sagst.
90. Verbesina perlanata Sagst. & Quipuscoa
91. Verbesina peruviana Sagst.
92. Verbesina piurana Sagst.
93. Verbesina plowmanii Sagst.
94. Verbesina sanchezii Sagst.
95. Verbesina simplicicaulis Sagst.
96. Verbesina subrotundifolia Sagst.
97. Verbesina weberbaueri Sagst.
Tabla 2. Continuacin.
Especies
1. Albizia multifora (Kunth) Barneby & J.W.Grimes var. sagasteguii Barneby & J.W. Grimes
2. Asplundianthus sagasteguii R.M. King & H. Rob.
3. Astragalus sagasteguii Gmez - Sosa
4. Byttneria sagasteguii Cristbal
5. Calceolaria sagasteguiana Rosencr.
6. Diplostephium sagasteguii Cuatrec.
7. Draba sagasteguii Al-Shehbaz
8. Epidendrum sagasteguii Hgsater & E. Santiago
9. Fleischmannia sagasteguii H. Rob.
10. Gentianella sagasteguii J.S. Pringle
Tabla 3. Taxones dedicados al doctor Abundio Sagstegui Alva. Compilado por Eric F. Rodrguez y Blanca Len.
(contina...)
134
Homenaje
Rev. peru. biol. 19(2): 127 - 134 (Agosto 2012)
11. Geranium sagasteguii Aedo
12. Gonolobus sagasteguii Morillo
13. Helianthopsis sagasteguii H. Rob.
14. Huperzia sagasteguiana B. llg.
15. Hydrocotyle sagasteguii Constance & M.O. Dillon
16. Jaltomata sagasteguii Mione
17. Koanophyllon sagasteguii R.M. King & H. Rob.
18. Larnax sagasteguii S. Leiva, Quipuscoa & N. W. Sawyer
19. Munnozia sagasteguii H. Rob.
20. Nasa sagasteguii Weigend
21. Ophryosporus sagasteguii H.Rob.
22. Passifora sagasteguii Skrabal & Weigend
23. Pentacalia sagasteguii H. Rob. & Cuatrec.
24. Piptochaetium sagasteguii Snchez Vega
25. Pitcairnia sagasteguii L.B. Sm. & Read
26. Puya sagasteguii L.B. Sm.
27. Rauhia sagasteguiana Ravenna
28. Tetramerium sagasteguianum T.F. Daniel
29. Tillandsia sagasteguii L.B. Sm.
30. Trixis sagasteguii Cabrera
31. Vernonia sagasteguii M.O. Dillon
32. Vriesea sagasteguii L.B. Sm.
Tabla 3. Continuacin.
135
La Revista basurero
Rev. peru. biol. 19(1): 135 - 138 (August 2012)
Rev. peru. biol. 19(1): 135 - 138 (Agosto 2012)
Facultad de Ciencias Biolgicas UNMSM
ISSN 1561-0837
La Revista basurero
Leonardo Romero
The Journal landfll
Facultad de Ciencias Biologicas, Universidad Nacional Mayor de San Marcos. Ciudad Universitaria de San Marcos Av. Venezuela s/n.
Apartado postal 110058, Lima 11, Per.
Email Leonardo Romero:
lromeroc@unmsm.edu.pe
EDITORIAL
Aumenta la investigacin y las publicaciones
cientfcas con todos sus males
El inters por publicar el resultado de nuestras investigaciones
es inherente a la investigacin, investigamos para descubrir la
verdad y comunicar lo que descubrimos. Sin embargo, al igual
que un ente vivo es modelado por las circunstancias o el ambiente
en que vive, la investigacin cientfca y sus publicaciones evo-
lucionan como consecuencia de cambios sociales, econmicos
y culturales. Los cambios sociales y en investigacin son tangi-
bles y pueden sintetizarse si se compara la informacin global
para el ao 2002 con el 2007
1
; el gasto global en investigacin
prcticamente se duplic de 790,3 billones en el 2002 a 1145,7
en el 2007, el nmero de investigadores se increment de 5,7
millones a 7,1; el nmero de publicaciones cientfcas aument
de 1,09 millones en el 2002 a 1,94 en el 2010, es decir en un
77%
1
. En estas diferencias, fueron las economas emergentes
como China, Brasil e India las que en conjunto aumentaron
sus inversiones en ciencia, tecnologa y desarrollo de 17 a 24%.
Aunque el Per todava no fgura en las estadsticas y ci-
fras generales de desarrollo de investigacin cientfca a nivel
mundial, toda una jerga sobre ciencia, tecnologa, desarrollo e
innovacin se escuchan en los pasadizos de nuestros institutos y
centros acadmicos, y responden a las intenciones de remontar
nuestro actual nivel cientfco (por lo menos el nivel en que nos
colocan bibliomtricamente
2
), dar a conocer nuestra produccin
en investigacin cientfca, y cmo ella puede ser la fuente de
desarrollo del pas. Para instituciones como la Universidad Na-
cional Mayor de San Marcos (UNMSM) y otras universidades
pblicas peruanas acreditar una consistente y robusta actividad
cientfca se convierte en un objetivo estratgico para deslindar
nuestra labor universitaria con la de otras instituciones seme-
jantes en la cuales el lucro es una de sus fnalidades principales.
Para la UNMSM, la acreditacin de su labor acadmica y de
investigacin, y el mostrar la calidad de su produccin intelec-
tual y cientfca sern los pilares que permitirn a la UNMSM
tener el peso necesario en las decisiones que involucren el
desarrollo nacional. Desde el ao 1998, en la UNMSM se
promovieron las revistas de investigacin, dentro del marco del
reglamento de investigaciones y como una herramienta para
facilitar la difusin de los resultados de las investigaciones en
las diferentes facultades. La Revista Peruana de Biologa es una
de esas revistas, pero que adems asumi el reto de convertirse
en un medio de excelencia para la publicacin de los trabajos
principalmente en las reas de biodiversidad y biotecnologa
en el Per. A pesar del tiempo transcurrido, su edicin an es
inestable, generalmente producido por problemas econmicos,
organizacionales y logsticos, causando la dilacin en el tiempo
de entrega como uno de sus problemas ms importantes. A estos
problemas se suman otros relacionados al quehacer de cualquier
revista cientfca incluso las del main stream, el problema de las
malas conductas y fraudes. Dentro del contexto de calidad que
la UNMSM busca, el incremento de trabajos fraudulentos se
torna un fenmeno delicado y a su vez demanda acciones ms
enrgicas por las consecuencias que pueden tener. Pero, rechazar
un trabajo fraudulento una vez detectado no es el problema, sino
la agobiante pregunta Por qu nos llegan tantos?
Qu no haran por publicar?
La publicacin de un artculo de investigacin cientfca
respalda la labor realizada, es uno de los productos ms impor-
tantes de las instituciones dedicadas a la ciencia y las labores
acadmicas y universitarias, crea y desarrolla el prestigio del
investigador y de la institucin ante la sociedad; todo esto aca-
rrea como consecuencia fondos para investigacin, bienestar,
crecimiento y desarrollo para el investigador y la institucin
(hechos no necesariamente vinculantes). Por otro lado, un editor
es un espectador y actor del drama de la publicacin cientfca
en el escenario del desarrollo de la ciencia. Publicar un trabajo
cientfco no es un tema sencillo y podra en algunos momentos
ser una actividad que se envuelva en una vorgine de intereses
de todo tipo, de colores, sabores, y malos olores. Uno de esos
momentos comienza cuando llega a la edicin un trabajo con
sntomas de malas conductas o fraudes. Aqu se enfrenta la
intencin del autor con la misin del editor que es de difundir
informacin de calidad y que satisfaga a la comunidad cientfca
y a la sociedad. Aunque estos malos comportamientos y fraudes
no son causados por la revista o el editor, sino provienen de la
comunidad cientfca misma, es el editor el que se bate con ellos
3
,
aunque ponga en riesgo su salud y su vida.
Para el Council of Science Editors
4
la mala conducta en in-
vestigacin se refere a malos tratamientos de los sujetos de
investigacin (personas o animales), la falsifcacin y fabricacin
de datos y la piratera y plagiarismo. Pero en otros mbitos se
separan los fraudes de las malas conductas por aspectos volitivos
y veniales. Mientras que, las malas conductas generalmente son
cometidas sin intensin, y pueden ser relativas a errores meto-
dolgicos (diseos experimentales, incumplimiento de normas
y recomendaciones), procedimientos culturales (como la falta
de tica en autoras, las publicaciones en salchichn); tenemos
por otra parte los fraudes, en los cuales la intensin del autor es
136
Romero
Rev. peru. biol. 19(1): 135 - 138 (Agosto 2012)
conscientemente expresa o velada pero innegable (falsear datos,
inventarlos, ocultar informacin, cocinar datos, plagio, etc.).
El lado oscuro de la publicacin cientfca
Oscuro porque el editor generalmente desconoce quin es el
autor y su trabajo, y ellos deben ser escudriados en su intensin
de publicar y en la veracidad de su informacin. Es decir en esta
primera escena de la revisin, los editores no sabremos de dnde
vendr el ataque o cmo vendr; en esa oscuridad se reciben los
trabajos. Aunque existen sistemas y software
5
para detectar el pla-
giarismo y otras malas conductas y fraudes, estos son un primer
tamiz en este aspecto. Sin embargo, varios editores consideran
a la primera revisin del trabajo como la parte del proceso en
que se cuentan con menos herramientas, personal adecuado y
tiempo para detectarlas
6
. Aunque parezcan demasiadas precau-
ciones, casi lindando con algn estado mental desquiciado, no
estamos muy lejos de las grandes editoras como Elsevier
7
que
ponen mucho nfasis en adiestrar a sus editores y autores sobre
estos temas, los aspectos legales para su incumplimiento y las
consecuencias de la infraccin.
La alegra de ser coautor sin responsabilidad
En este drama identifcar las malas conductas y fraudes es
uno de los papeles del editor y de los revisores (peer review). Sin
embargo, los actuales modos de investigacin interdisciplinarios,
las investigaciones en grandes laboratorios, y donde es frecuente
que se produzcan artculos con muchos autores, crean una escena
en la que el responsable del artculo puede cargar con la culpa de
la mala conducta y fraude
8
(en muchos casos con sanciones de
parte de su institucin) y los otros autores (mal denominados,
pero convenientemente coautores) no sufren ms sanciones que
la vergenza. Y la pregunta es donde surgi el problema, no
hubo discusin entre los autores? No se compartieron datos
entre autores? Cmo interactuaron? Cmo se hizo el trabajo?
Todos no tienen las mismas responsabilidades? Este aspecto nos
lleva a la conclusin de que las instituciones, y no solamente los
miembros de los comits editoriales, deben asumir e identifcarse
con las buenas prcticas de investigacin y buena conducta, de
manera que la conducta fraudulenta pueda ser por lo menos
identifcada y aislada
9
. La participacin de todos los autores en
la presentacin del trabajo, la posibilidad de contactarse con cada
uno de los autores, cartas de compromiso y otros, son fltros que
estn dirigidos a identifcar una posible mala conducta.
El peer review como fltro
El revisor de un trabajo que pretende ser publicado es un par
(o igual) o muchas veces otro cientfco con ms experiencia en
el tema y que puede dar luces como mejorar la presentacin de
la informacin. El revisor (peer review) es una instancia presente
en la edicin cientfca casi desde los inicios de la publicacin
cientfca hace ms de 300 aos. Una encuesta realizada a autores
de revistas de alto factor de impacto (FI) en el ao 2008
10
a pe-
dido del Publishing Research Consortium, mostr que el 43% de
los encuestados consideraba que el peer review fltra los trabajos
fraudulentos y con malas conductas. Ante esta percepcin se
enfrentan investigaciones que ponen en tela de juicio la severidad
y consistencia del peer review para detectar casos de fraude
11
.
Los revisores juegan un papel importante en la determinacin
de la calidad del trabajo, la pertinencia y la actualidad, por su
experiencia son los que estn en mejores condiciones de detec-
tar mala conducta. Sin embargo, la fabricacin o invencin de
resultados es ms difcil de detectar sobre todo en determinados
temas en los cuales se arguye la imposibilidad de la repeticin del
experimento. Pero en el rea de biodiversidad, detectar especies
que no existen en el lugar, o especies inventadas es ms fcil
por la experticia de los revisores. Por lo tanto el revisor es un
fltro fno, dedicado a ciertos tipos de malos comportamientos
y seleccionar trabajos de calidad y con informacin relevante.
La retraccin, la punta del iceberg
Los niveles en que el plagiarismo y las malas conductas son
muy altos. En el 2010 la editora Francis & Taylor del grupo
Nature Publishing escudrio los trabajos recibidos en los ltimos
meses y en una de sus revista se detect el 23% de los trabajos
con algn grado de plagiarismo
12
. Recientemente Daniele Fanelli
analizando los artculos que solo han perpetrado fraudes como
falsifcacin, fabricacin de datos, alteracin de informacin,
etc. encuentra que casi el 2% admite haber cometido fraude y
el 34% otro tipo de mala conducta grave
13
. En general esto es
considerado como la punta del iceberg porque muchos trabajos
no pasan exmenes tan minuciosos.
La retraccin es la accin de retirar un artculo de circulacin,
se publica y comunican los motivos por los que el artculo no
debe ser citado ni mencionado como referencia. Lo cual es
considerado como un acto muy vergonzoso para los autores y
su institucin, adems de ser un trabajo adicional al proceso de
edicin. En octubre 2012, Fang et al.
14
publicaron en PNAS un
artculo que ha sido comentado y citado en varios peridicos y
algunas revistas. Ellos analizaron poco ms de 2000 artculos de
la base de datos de PubMed y encontraron que el 43% fueron
retractados por ser fraudes y el 14 y 10% fueron por publicacin
duplicada y plagiarismo respectivamente. Algunos investigadores
pueden ver a la retraccin como una penalidad inocua y sin
importancia, sin embargo existen observadores como Retrac-
tion Watch (http://retractionwatch.wordpress.com) que es un
blog donde se documentan estos casos ; y podemos mencionar
el caso de Scott Reuben que tuvo varios trabajos retractados en
la revista Anesthesiology News y que posteriormente fue enjui-
ciado por fraude de atencin mdica, condenado a seis meses
de prisin federal y el pago de $ 415000 en multas, decomiso
y la restitucin
15
.
Cokol
16
et al. ensayaron un modelo para tener una idea de
cuantos trabajos deberan de ser retractados. Describe un modelo
estocstico con parmetros que implican el rigor para aceptar
una pequea fraccin de "manuscritos fallados" y dos factores
sobre el escrutinio post-publicacin en funcin del factor de
impacto (IF). Entre sus conclusiones encuentra que las revistas
con revisiones exigentes tanto las de alto IF como las de bajo
IF presentaran menos artculos que podran ser retractados.
Adems, en aquellos con ms alto IF, la retraccin es ms severa
en post-publicacin. Aunque sus resultados relacionaran la
efciencia de la produccin con las probabilidades de publicar
artculos con mala conducta; tambin podramos interpretar
que los autores inescrupulosos identifcan aquellas revistas que
son ms permisivas, y que no tienen fltros exigentes sobre los
trabajos fraudulentos o con comportamientos de mala conducta.
Evidentemente si se tratan de comportamientos volitivos, de
los cuales se conocen sus consecuencias, tendremos un modelo
donde la tendencia es a atiborrar con trabajos indeseables a las
revistas con menos recursos.
137
La Revista basurero
Rev. peru. biol. 19(1): 135 - 138 (August 2012)
Quiero publicar mi trabajo
Algo de lo mencionado arriba suceder en la Revista Pe-
ruana de Biologa? La respuesta es s. Por lo menos la llegada
de trabajos fraudulentos y con malas conductas s suceden y
frecuentemente. Es una experiencia que se puede comprobar, si
alguien desea obtener trabajos fraudulentos lo nico que tiene
que hacer es emprender una revista cientfca, llevar a cabo una
revisin por pares, lograr visibilidad en bases de datos, ser ms
o menos constante y esperar, pronto llegaran.
Qu esta sucediendo? Simplemente que los editores de
revistas pequeas se enfrentan no solo a la problemtica de la
investigacin nacional, a la falta de recursos, polticas de inves-
tigaciones dbiles o inexistentes, sino al maremgnum de inves-
tigadores que quieren que sus investigaciones sean publicadas,
generalmente presionados por compromisos de diversa ndole
y no porque ellos piensen que la informacin en su trabajo es
relevante o contribuir con la ciencia. Entonces, ser comn ver
como son sometidos a la Rev peru biol los trabajos que fueron
rechazados en otras revistas, y las que rechacemos irn a otras,
en una cascada hasta que por fn lleguen a una revista donde
sern publicados. La revista que se ubique al fnal de la cascada
ser una revista basurero.
Si bien es cierto en la Rev peru biol hemos recibido trabajos
fabricados, plagios, auto-plagios, trabajos fragmentados; los ms
comunes son aquellos en que se omite informacin (consciente
o inconscientemente). Algunas veces sobre el mismo trabajo,
ocultando errores de muestreo o por ejemplo un experimento
donde ms de la mitad de individuos de prueba murieron por
razones desconocidas antes de culminar el experimento. Otras
omiten informacin sobre trabajos similares que deberan de
discutirse, en algunos casos por no tener acceso a la publicacin
original, otras porque sus resultados no se pueden contrastar,
mostrando fallas metodolgicas, o por que pondran en evidencia
otro tipo de errores, incluso conceptuales. Otro tipo frecuente
es la fragmentacin de los trabajos, donde en lugar de brindar
un trabajo completo que de luces a un problema, muestran
parte por parte la informacin sin un trabajo de anlisis y
aporte signifcativo, as por ejemplo como si quisiramos dar a
conocer inventarios de ambos lados de una quebrada en sendos
artculos. La publicacin de todos estos tipos de trabajos sola-
mente es contraproducente y daina no solo para la revista sino
para la institucin y los cientfcos nacionales en general por el
descredito que sufriran nuestras investigaciones.
Este fenmeno debe ser enfrentado dentro de cada una de las
revistas que se proponga llegar a la calidad que tanto esperan sus
usuarios y tambin por las instituciones que apuestan (e invierten
muchas veces sus escasos recursos) por realizar investigacin
cientfca como la UNMSM. Debe dedicarse considerable
esfuerzo para no publicar trabajos fraudulentos, por los obvios
motivos de mantener la reputacin, la credibilidad, y el peso
de nuestras ideas y propuestas en la sociedad. La realizacin de
este esfuerzo disminuir la llegada de esos tipos de trabajos. Pero
tambin, debemos aplicar la censura post-publicacin, y debe
ser asumida por la institucin y practicada por las revistas. Las
retracciones deben de ser indicadores de nuestro esfuerzo de
mejorar nuestra publicacin, de lograr la credibilidad y calidad.
La institucin debe internalizar en sus polticas los aspectos
de evaluacin de calidad del trabajo de investigacin y de las
publicaciones y no solamente de los procedimientos y normas.
Una revista sola no podr sobrevivir en este escenario, para
ella el drama acabara en tragedia; la trama empezara cuando
ella comience a incrementar su esfuerzo en prevenir el fraude,
pronto la cantidad de artculos comenzar a disminuir (no es
una revista fcil, pensaran los autores inescrupulosos), luego
no podr llegar a los estndares de publicacin de las bases de
datos ms importantes (que piden ms artculos por ao) y lu-
ego se extinguir lentamente. Para sobrevivir necesitar de una
comunidad cientfca que la apoye, una comunidad crtica que
tenga clara su misin con respecto de la ciencia y la sociedad.
Necesitar de instituciones con polticas coherentes y robustas,
que se fjen los objetivos a alcanzar.
Referencias
1 Te Royal Society. 2011. Knowledge, networks and nations: Global scientifc
collaboration in the 21st century. RS Policy document 03/11. London.
http://royalsociety.org/policy/reports/knowledge-networks-nations.
2 Pea V. A., Rivera P., Schulz P.A.B. & Tapia J.R.B. 2011. La Produccin
Cientfca en San Marcos. Hechos, cifras y estndares internacionales
2002-2010. Ediciones Vicerrectorado Acadmico UNMSM. CONCY-
TEC. 684pp.
3 Marusic A., V. Katavic & M. Marusic. 2007. Role of editors and journals in
detecting and preventing scientifc misconduct: strengths, weaknesses,
opportunities, and threats. Medicine and law 26 (3): 545566.
4 Scott-Lichter D. & the Editorial Policy Committee Council of Science
Editors. 2012. CSEs White Paper on Promoting Integrity in Scientifc
Journal Publications, 2012 Update. 3rd Revised Edition. Wheat Ridge,
CO: Available at: http://www.councilscienceeditors.org/fles/public/
entire_whitepaper.pdf.
5 http://www.ithenticate.com/resources/overview/
5 http://www.crossref.org/crosscheck/index.html
6 DeMaria A.N. 2012. Scientific Misconduct, Retractions, and Errata.
Journal of the American College of Cardiology 59 (16): 14881489.
doi:10.1016/j.jacc.2012.03.005.
7 http://www.elsevier.com/about/publishing-guidelines/publishing-ethics
8 Franzen M., S. Rdder & P. Weingart. 2007. Fraud: causes and culprits as
perceived by science and the media. Institutional changes, rather than
individual motivations, encourage misconduct. EMBO Reports 8 (1):
37. doi:10.1038/sj.embor.7400884.
9 Kreutzberg G.W. 2004. Te rules of good science. EMBO reports 5 (4):
330332. doi:10.1038/sj.embor.7400136.
138
Romero
Rev. peru. biol. 19(1): 135 - 138 (Agosto 2012)
10 Publishing Research Consortium. 2008. Peer review: benefts, perceptions
and alternatives. Mark Ware Consulting. http://www.publishingresearch.
net/documents/PRCsummary4Warefnal.pdf
11 Peters D.P. & S.J. Ceci. 1982. Peer-review practices of psychological journals:
Te fate of published articles. Behavioral and Brain Sciences 5 (2):
187255. doi:10.1017/S0140525X00011183.
12 Butler, D. 2010. Journals step up plagiarism policing. Scientifc American.
http://www.scientificamerican.com/article.cfm?id=journals-police-
plagiarism
13 Fanelli D. 2009. How Many Scientists Fabricate and Falsify Research? A
Systematic Review and Meta-Analysis of Survey Data. PLoS ONE 4
(5): e5738. doi:10.1371/journal.pone.0005738.
14 Fang F.C., R.G. Steen & A. Casadevall. 2012. Misconduct Accounts for
the Majority of Retracted Scientifc Publications. Proceedings of the
National Academy of Sciences 109 (42): 1702817033. doi:10.1073/
pnas.1212247109.
15 O'Reilly K.B. 2012. Misconduct accounts for most journal retractions. AMA.
Amednews.com. http://www.ama-assn.org/amednews/2012/10/15/
prsb1015.htm
16 Cokol M., I. Iossifov, R. Rodriguez-Esteban & A. Rzhetsky. 2007. How
Many Scientifc Papers Should Be Retracted? EMBO Reports 8 (5):
422423. doi:10.1038/sj.embor.7400970.
139
Una nueva especie de Salvia del Norte de Per
Rev. peru. biol. 19(2): 139 - 142 (August 2012)
Rev. peru. biol. 19(2): 139 - 142 (Agosto 2012)
Facultad de Ciencias Biolgicas UNMSM
ISSN 1561-0837
Una nueva especie de Salvia (Lamiaceae) del Norte de Per
Abundio Sagstegui Alva
1
y Eric F. Rodrguez Rodrguez
2
A new species of Salvia (Lamiaceae) from Northern Peru
1 Herbarium Truxillense (HUT),
Universidad Nacional de Trujillo.
Jr. San Martn 392. Trujillo, Per.
2 Herbarium Truxillense (HUT),
Universidad Nacional de Trujillo.
Jr. San Martn 392. Trujillo, Per.
Email Eric F. Rodrguez Rodrguez:
efrr@unitru.edu.pe
Presentado: 07/05/2012
Aceptado: 18/08/2012
Publicado online: 10/11/2012
Resumen
Se describe e ilustra una nueva especie de Salvia L. (Lamiaceae), denominada Salvia vargas-llosae Sagst.
& E. Rodr. sp. nov. perteneciente a la seccin Cylindriforae (Epling) Epling, procedente de los andes del Norte
del Per (provincia Celendn, departamento Cajamarca), tpica de la jalca Sendamal-Kumulca-Challuayaco
sobre los 3000 m de altitud y aparentemente endmica a esta parte del pas. Se discute con sus relacionadas
y adicionalmente se presentan datos sobre su distribucin geogrfca, ecolgica y estado de conservacin.
Palabras clave: Salvia, Cylindriforae, Lamiaceae, especie nueva, Norte del Per.
Abstract
Salvia vargas-llosae Sagst. & E. Rodr. sp. nov. is described and illustrated as a new species of Salvia L.
(Lamiaceae) from the Andes of Northern Peru (province of Celendin, department of Cajamarca). This new spe-
cies belongs to the section Cylindriforae (Epling) Epling and typically restricted to the region jalca (Sendamal-
Kumulca-Challuayaco) above the 3000 m of altitude and apparently endemic to this part of the country. It is
discussed with its closest relatives and additionally data on its geographical distribution, ecology and state of
conservation are provided.
Key words: Salvia, Cylindriforae, Lamiaceae, new species, Northern Peru.
Introduccin
El gnero cosmopolita Salvia L., el ms extenso de la familia
Lamiaceae (=Labiatae), cuenta en la actualidad con unas 935
especies reconocidas (Fernndez-Alonso 2008 a, b). Para la fora
del Per se han registrado 79 taxones entre especies y subespecies,
siendo catalogado como el ms diverso de la familia (Zarucchi
1993, Granda 2010). Actualmente, son consideradas a 32
especies endmicas, constituyndose en el gnero ms rico en
endemismos de la familia (Rodrguez 2006, Granda 2010). Los
ltimos estudios que se han efectuado con material peruano son
los realizados por Epling & Jtiva (1963, 1966, 1968) y Granda
(2010). La mayora de especies presentes en el Per se encuentran
registradas para el norte. Especialmente los andes constituyen un
importante hbitat. Revisando crticamente tanto el material de
herbario como el de las ltimas colecciones procedentes de los
andes peruanos, en particular del Norte, se ha encontrado una
nueva especie que denominamos Salvia vargas-llosae Sagst. &
E. Rodr. sp.nov. procedente de la cadena central de los andes
del norte en la zona de Cajamarca, Prov. Celendn. Se debe
enfatizar que de esta cadena central nortea, perteneciente a la
zona de Amotape-Huancabamba de elevada riqueza endmica
(Weigend 2002, 2004), se han descrito nuevos taxones para
la ciencia de material recolectado por personal del Herbarium
Truxillense (HUT) de la Universidad Nacional de Trujillo (e.g.:
Salvia lanicaulis Epling & Jtiva, S. medusa Epling & Jtiva,
Salvia xanthophylla Epling & Jtiva) (Epling & Jtiva 1963).
Consecuentemente, la descripcin, ilustracin y discusin de
este nuevo taxn es el objetivo de este trabajo.
Material y mtodos
El estudio est basado en la revisin de material de herbario
(CPUN, HUT, F, MO) y observaciones directas de hbito y
hbitat en el campo. Las colecciones se realizaron de acuerdo
con la metodologa y tcnicas convencionales de herborizacin
(Rodrguez & Rojas 2002). Adicionalmente al trabajo de campo
se fj y conserv material en lquido (alcohol etlico al 70%
o AFA) para estudiar la estructura foral. El material botnico
del tipo fue depositado en los siguientes herbarios: HUT, F,
MO. Son presentadas, la descripcin, discusin, delineacin
y mediciones de la especie (Fig. 1 A-H). Se adicionan datos
sobre su distribucin geogrfca, ecologa, fenologa y estado
de conservacin segn criterios de la Lista Roja UICN (UICN
2001). Los acrnimos de los herbarios son citados segn Hol-
mgren et al. (1990).
Taxonoma
Salvia vargas-llosae Sagst. & E. Rodr. sp.nov.
(Fig. 1 A-H)
Tipo: PER. Dpto. Cajamarca, Provincia Celendn, Cha-
lluayaco (La Encaada-Celendn), ladera rocosa, 3600 m, 18
mayo 1976, A. Sagstegui A., J. Cabanillas S. & O. Dios C. 8409
(Holtipo: HUT-13705; Istipos: F, MO).
Herba perenne, ca. 50 cm alta, erecta vel ascendens, parce ra-
mosa, rhizomatosa. Caulis tetragonis, 2-3 mm crassis, pilis septatis
aglandularis, ca. 1 mm longis et pilis septatis glandularis, ca. 0,5
mm longis. Folia opposita, petiolata; petiolo acanalato; lamina
ovata, basi cordata, apice obtusa, crenata, 16-22 14-20 mm.
Inforescentiae racemosae terminales, (5-) 7-15 (-17), verticillastrae
4-7(-8) forae. Bracteis late ovato-ellipticis, acuminatis, caducis,
fuscescentibus, externe pilosis. Calyx bilabiatis, 10-13 5-6 mm;
superne labium 4-6 3-3,5 mm, 6-nervatis, inferne labium
2-lobatum, lobulis breviter apiculatis, 4-5 2-3 mm, 3-nervatis.
Corolla violacea, tubo 18-20 mm longo, superne labium 7-8 mm
longum, inferne labium 12-15 mm longum, 3-lobulatum, latere
lobulis ellipticis, 2-3 2 mm, mdium 2-lobulatis, 5-6 6-8 mm.
Filamentum adnatum ca. 15 mm ad basim tubular; antherae ba-
sifjae. Ovarium ellipsoideum, 2 1,2 mm; stylis bifdus. Nucula
ellipsoide, fuscescentibus, maculata, 2,5-3 1,5 mm.
140
Sagstegui y Rodrguez
Rev. peru. biol. 19(2): 139 - 142 (Agosto 2012)
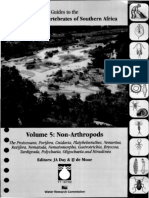
Figura 1. Salvia vargas-llosae Sagst. & E. Rodr. A. Rama forfera; B. Hoja; C. Flor; D. Brctea de la parte media de la inforescencia; E.
Estambres mostrando al flamento, conectivo y anteras; F. Ovario, estilo y estigmas; G. Disposicin de las nculas; H. Ncula. Delineado por
R. Aguirre T. del holtipo: A. Sagstegui A., J. Cabanillas S. & O. Dios C. 8409 (HUT).
141
Una nueva especie de Salvia del Norte de Per
Rev. peru. biol. 19(2): 139 - 142 (August 2012)
Hierba perenne de hasta 50 cm de altura, erecta, decumbente
o ascendente, estolonfera, poco ramifcada, solamente en la parte
inferior, ramitas hasta 7 cm de largo; indumento blanquecino
piloso-glanduloso, con pelos simples septados aglandulares, 4-7
clulas, clula basal buliforme, de hasta 1 mm de largo y pelos
simples septados con pice glandular, 3-4 clulas, de hasta 0,5
mm de largo. Tallos de 2-3 mm de dimetro, tetrgonos, gene-
ralmente rojizos, con indumento concentrado en la zona media
de las caras y escaso en las aristas, constituido mayormente por
pelos glandulares; entrenudos 10-65 (-75) mm de longitud.
Hojas opuestas, pecioladas (pecolo canalado, 10-15 mm de
longitud, pubescencia abundante constituida principalmente
por pelos simples aglandulares), lminas ovadas, pice obtuso,
base cordada, crenadas (dientes subredondeados e irregulares,
10-12 a cada lado), con 3-4 nervios secundarios a cada lado
del nervio medio, nervios de tercer y cuarto orden poligonal,
nerviacin poco resaltada en el envs; haz verde oscuro y envs
verde claro, pubescentes en la haz, excepto en las nervaduras,
indumento constituido de pelos simples septados aglandulares,
igual indumento pero ms corto y disperso en las nervaduras
del envs; de 16-22 14-20 mm en las mas grandes. Info-
rescencia terminal, racemosa, interrumpida, 80-180 mm de
largo, constituida por (5-) 7-15 (-17) verticilastros, 4-7(-8)
fores (antesis asincrnica), separados 5-40 mm entre s (menor
separacin hacia el pice), parte basal del raquis sin fores de 45-
110 mm, densamente hrtulo, con indumento generalmente no
glandular; brcteas ampliamente ovado-elpticas, acuminadas,
caducas, marrones, exteriormente pubescentes, dominancia de
pelos simples no glandulares sobre los glandulares, 8,5-9 5,5-
6 mm; pedicelos de 5-6 mm de longitud en la antesis y 8-11
mm en la fructifcacin, cortamente hrtulos, con indumento
simple septado no glandular. Cliz bilabiado, de 10-13 mm de
longitud y 5-6 mm de ancho en la antesis, 14-18 6-7 mm en
fructifcacin, tubo 7-10 mm de largo, externamente con in-
dumento corto, hirto, dispuesto en los nervios, predominancia
de pelos glandulares uniformemente distribuidos, internamente
glabros; labios muy abiertos, desiguales, el superior de 4-6 mm
de longitud y 3-3,5 mm de ancho, con 6 nervios conspicuos de
color morado intenso, el inferior bidentado, dientes brevemente
apiculados, de 4-5 2-3 mm, con 3 nervios conspicuos verdes
cada uno. Corola violcea, indumento abundante formado por
pelos simples no glandulares septados lanuginosos, violceos,
de hasta 0,5 mm de longitud; tubo de la corola de 18-20 mm,
recto y gradualmente ensanchado desde la base (3-3,5 mm de
anchura) hasta el pice (6 mm de anchura); labio superior de
7-8 mm de longitud, labio inferior amplio, patente, 12-15 mm
de largo, 3-lobulado, lbulos laterales elpticos, 2-3 2 mm,
lbulo medio ms amplio, escotado, 2-lobulado, 5-6 6-8 mm,
sobrepasando ca. 2-3 mm en longitud a los laterales. Estambres
con flamentos adnatos a ca. 15 mm de la base del tubo, rojizos,
glabros, de 4 mm de largo; conectivo levemente arqueado, ro-
jizo, de 20 mm de longitud, sobrepasando 10-12 mm al labio
superior; anteras estrechamente cilndricas, basifjas, oscuras,
polen amarillo, de 3 mm. Ovario elipsoidal, amarillento, 2
1,2 mm; estilo de 35-40 mm de longitud, rojizo, glabro, con
ramas estigmticas desiguales, la superior de 3 mm y la inferior
de 1-1,5 mm, divergentes, claramente ensanchado-espatulada
y obtusa en el extremo distal. Nculas elipsoidales, de color
marrn claro con mculas marrn oscuro y contornos redon-
deados, de 2,5-3 1,5 mm.
Material Adicional Examinado:
PERU: Dpto. Cajamarca. Prov. Celendn: Chaluayaco,
entre Kumulca y Celendn, 3350 m, 4 Mayo 1970, I. Sn-
chez V. & W. Ruiz V. 469 (CPUN-3192). En el lugar llamado
Challuayaco, sobre la carretera a Celendn, al este del paso de
Cumullca, 3550 m, 12 Mayo 1984, I. Snchez V. & W. Ruiz V.
3474 (CPUN-3140). Distr. Encaada, Cumulca, 7 Abril 1966,
I. Snchez V. & M. Alva Z.189 (CPUN-3166). Sendamal, entre
la Encaada y Celendn, 3330 m, 3 Julio 1975, I. Snchez V. &
W. Ruiz V. & J. Snchez V. 1691 (CPUN-1691). Challuayaco
(Encaada-Celendn), 3350 m, 4 Mayo 1970, A. Sagstegui A.
7388 (HUT-7402). Jalca de Kumulca, 3500 m, 01 Julio 1978,
A. Lpez M. & A. Aldave P. 8452 (HUT-15381).
Discusin Taxonmica:
Salvia vargas-llosae Sagst. & E. Rodr. sp.nov. se encuentra
circunscrita en la Seccin Cylindriforae (Epling) Epling prin-
cipalmente porque es una hierba perenne tpica de los Andes
con 4-7(-8) fores en cada verticilastro, corola desde rojiza hasta
violcea, parte interna del tubo de la corola sin indumento ni
papilas, estambres bien exertos y adnatos sobre la mitad del tubo
de la corola, as como por presentar estilos glabros. De todas
las especies de la Seccin con la que presenta mayor afnidad
es con S. ayavacensis Kunth (= Salvia mucidistachys Epling)
(ver Bonpland & Humboldt 1817, Macbride 1960, Zarucchi
1993), por el tipo de indumento, forma y margen de las hojas,
nmero de fores por verticilastro, color de fores, tamao del
flamento del estambre, entre otras caractersticas. Sin embar-
go, S. ayavacensis se diferencia claramente de la nueva especie
porque es una planta robusta (mayor a 1 m versus hasta 50 cm
de alto), hojas mas grandes y pecolos mas largos, disposicin
de los verticilastros en la inforescencia, fores ms grandes (e.g.:
cliz de 17 mm versus 10-13 mm de largo en antesis, tubo de la
corola ca. 3 cm versus 2 cm de largo, conectivo del estambre ca.
40 mm de largo versus 20 mm de largo en la especie nueva), y
pedicelos forales en antesis son ca. 2 veces y media mas largos a
los ocurridos en S. vargas-llosae (15-16 mm versus 5-6 mm). As
mismo, S. ayavacensis es una especie endmica al departamento
de Piura (Rodrguez 2006) [cerca al ro Cutaco, prov. Ayavaca-
tipo, y Talaneo-Jalca Cixse, prov. Huancabamba, ver coleccin:
A. Sagastegui A. & J. Cabanillas S. 8634 (HUT-13961)] y que
crece en los Andes de la vertiente occidental nortea a altitudes
ms bajas (bajo los 3,000 m de altitud)en relacin con la especie
nueva. En general S. vargas-llosae es la especie que presenta las
fores ms pequeas de la Seccin. Se debe precisar que con Salvia
lanicaulis Epling & Jtiva [ver colecciones de jalca Kumulca:
A. Sagstegui A., J. Mostacero L. & S. Leiva G. 12039 (HUT-
19712), A. Sagstegui A., J. Cabanillas S. & O. Dios C. 8113
(HUT-13210), otra especie de la misma Seccin, comparten
solamente la distribucin geogrfca y hbitat (Jalca) a mas de
3000 m. As mismo, otras especies norteas pertenecientes a la
misma Seccin, como S. cylindrifora Epling y S. hirta Kunth
de Piura, y S. heerii Regel de Cajamarca, crecen en diferentes
hbitats y a altitudes mucho ms bajas, pero mayormente carecen
de afnidad con el nuevo taxn.
Distribucin, Ecologa y Fenologa:
Salvia vargas-llosae es aparentemente endmica de la jalca
de la Provincia Celendn, Cajamarca (localidades de Sendamal,
Kumulca y Challuayaco), distribuyndose arriba de los 3,000
142
Sagstegui y Rodrguez
Rev. peru. biol. 19(2): 139 - 142 (Agosto 2012)
m de altitud. Es una de las pocas especies andinas de Salvia que
crece en altitudes elevadas. En las localidades mencionadas es
encontrada con relativa frecuencia formando poblaciones de
pocos individuos. Habita laderas pedregosas expuestas, suelos
negros de humifcacin variable, musgosos, as como en am-
bientes con humedad edfca; asociada con hierbas, arbustos y
vegetacin propia del rea que incluye a Salvia lanicaulis Epling
& Jtiva. La nueva especie est restringida a la denominada
zona de Amotape-Huancabamba de elevada riqueza endmica
(Weigend 2002, 2004).
Florece y fructifca en mayo despus de las lluvias. Es visitada
por colibres e insectos (planta melfera).
Etimologa:
La nominacin de este taxn nuevo para la ciencia la aso-
ciamos con el Premio Nobel de Literatura 2010 el peruano Dr.
Mario Vargas Llosa para felicitarlo y adherirnos a su prestigio
universal, a quien rogamos acepte esta forma de hacerlo en ca-
lidad de Acadmicos y Cientfcos de la Universidad Nacional
de Trujillo, Per, en donde por felicidad nos ocupa la Scientia
Amabilis, es decir el estudio de las plantas que son los seres que
determinan la vida en el mundo.
Estado de Conservacin:
La nueva entidad hasta el momento es considerada endmica
al norte del Per y actualmente su distribucin se encuentra
restringida a la jalca de la provincia Celendn, Cajamarca. Se-
gn las colecciones efectuadas generalmente en el mismo lugar,
el rea de presencia y ocupacin es relativamente pequea, el
nmero y tamao de sus poblaciones son pequeas y dispersas.
Adems, la preocupacin esta focalizada en que esta especie vive
en ecosistemas frgiles que no estn protegidos por el Estado
peruano y en donde es notoria la prdida y alteracin progresiva
de hbitat (e.g.: quemas peridicas, pastoreo, cultivo de conferas
como Pinus radiata D. Don.). De acuerdo con esta informacin
y segn los criterios de la Lista Roja UICN (UICN, 2001),
debera ser incluida en la categora VU (Vulnerable).
Agradecimientos
Agradecemos a nuestro maestro Dr. Arnaldo Lpez Miranda
(HUT) por sus enseanzas y dirigir los trabajos de campo en
el Norte del Per; a la Dra. Nancy Hensold (F) por advertir la
presencia de esta nueva entidad; al Dr. Isidoro Snchez Vega
(CPUN) por el prstamo de material adicional; al Dr. Jim So-
lomon (MO) por confrmar la presencia del tipo en el herbario
MO; y a la Biloga Roxana Aguirre por la preparacin de la
excelente ilustracin que forma parte de esta investigacin.
Literatura citada
Bonpland A. & A. de Humboldt. 1817. Nova Genera et Especies
Plantarum. Tomo II. Lutetiae Parisiorum, Sumtibus
Librariae Greco-Latino-Germanice, via dicta Fosses-
Montmartre, No 14.
Epling C. & C. Jativa. 1963. Supplementary notes on American
Labiatae. VIII. Brittonia 15(4): 366-376.
Epling C. & C. Jativa. 1966. Supplementary notes on American
Labiatae. IX. Brittonia 18(3): 255-265.
Epling C. & C. Jativa. 1968. Supplementary notes on American
Labiatae. X. Brittonia 20(4): 295-313.
Fernndez-Alonso J.L. 2008a. Estudios en Labiatae de Colombia
VII. Salvia yukoyukparum, nueva especie y primer repre-
sentante de la Seccin Tomentellae en Colombia. Novon
18: 38-42.
Fernndez-Alonso J.L. 2008b. Estudios en Labiatae - VI. Hibridacin
en el gnero Salvia en Colombia y su inters horticultural.
Caldasia.30 (1):.21-48.
Granda A. 2010. Salvia hunzikeri (Lamiaceae), una nueva especie
de los Andes del Per. Rev. peru. biol. 17(2): 151-154.
Holmgren P. K., N. H. Holmgren & L. C. Barnett. 1990. Index Her-
bariorum. Part. I: The Herbaria of the World. 8
th
. ed. The
New York Botanical Garden, Bronx, New York. U.S.A.
Macbride F. 1960. Labiatae. En Flora of Peru. Field Museum of
Natural History, Botany 13(5/2): 721-829.
Rodrguez E. & R. Rojas. 2002. El Herbario: Administracin y
Manejo de Colecciones Botnicas. Edit. por R. Vsquez
M., Missouri Botanical Garden, St. Louis, U.S.A.
Rodrguez M. 2006. Lamiaceae endmicas del Per. En El libro rojo
de las plantas endmicas del Per. Ed.: B. Len et al. Rev.
peru. biol. Nmero especial 13(2): 371-379.
UICN. 2001. Categoras y Criterios de la Lista Roja de la UICN.
Versin 3.1. Preparado por la Comisin de Supervivencia
de Especies de la UICN. UICN, Gland, Suiza y Cambridge,
Reino Unido.
Weigend M. 2002. Observations on the Biogeography of the
Amotape-Huancabamba Zone in Northern Peru. In: K.
Young et al., Plant Evolution and Endemism in Andean
South America. Bot. Review 68(1): 38-54.
Weigend M. 2004. Additional observations on the biogeography
of the Amotape-Huancabamba zone in Northern Peru:
Defning the South-Eastern limits. Rev. peru. biol. 11(2):
127-134.
Zarucchi J. L. 1993. Lamiaceae. In: Brako, L. & Zarucchi, J.L.
(eds.). Catlogo de las Angiospermas y Gimnospermas del
Per. Monographs in Systematic Botany from the Missouri
Botanical Garden 45: 579-590.
143
Nueva especie de Deprea (Solanaceae) del Per
Rev. peru. biol. 19(2): 143 - 147 (August 2012)
Rev. peru. biol. 19(2): 143 - 147 (Agosto 2012)
Facultad de Ciencias Biolgicas UNMSM
ISSN 1561-0837
Una nueva especie de Deprea Raf. (Solanaceae) del Per
Marco Antonio Cueva
e talo Francisco Trevio
A new species of Deprea Raf. (Solanaceae) from Peru
Herbarium Arequipense (HUSA),
Universidad Nacional de San Agus-
tn de Arequipa, Av. Daniel Alcides
Carrin s/n.
Email Marco A. Cueva:
mcueva5@hotmail.com
Email Italo F. Trevio:
ifrant01@gmail.com
Presentado: 16/05/2012
Aceptado: 08/10/2012
Publicado online: 10/11/2012
Resumen
Describimos la nueva especie Deprea oxapampensis M. Cueva & I. Trevio (Solanaceae) a partir de un ejem-
plar de Oxapampa (Pasco, Per). Se caracteriza por tener hojas elpticas de 6,57,8 1,92,8 cm, 36 fores
axilares, pedicelos de 1119 mm de longitud, corola anaranjada tubuloso-urceolada con solo 5 lbulos, cliz
de 2,7 mm de longitud y fruto elipsoidal con el cliz acrescente adpreso a la baya. Se discute adems sobre
sus relaciones con D. ecuatoriana Hunz. & Barboza.
Palabras clave: Deprea; especie nueva; Solanaceae; Pasco; Per.
Abstract
Deprea oxapampensis M. Cueva & I. Trevio (Solanaceae) is described from a specimen collected in Oxa-
pampa (Dpto. Pasco, Peru). The new species is characterized by elliptical leaves 6,57,8 1,92,8 cm, 36
axillary fowers, pedicels 11-19 mm long, orange tubular-urceolate corolla with only 5 lobes, calyx 2,7 mm long
and elliptical fruit with acrescent calyx adpressed to the berry. We discuss about its relationships with D. ecu-
atoriana Hunz. & Barboza.
Keywords: Deprea; new species; Solanaceae; Pasco; Peru.
Introduccin
Deprea es un miembro de la subfamilia Solanoideae, y est
incluida dentro de la tribu Solaneae (Hunziker 2001). Segn
Barboza & Hunziker (1994) las especies de este gnero fueron
relacionadas con las de otros gneros de la tribu como Physalis,
Withania, Athenaea y Larnax, siendo ste ltimo considerado
como el ms afn.
Existen varias caractersticas morfolgicas a nivel de la for
que distinguen notoriamente a Deprea de Larnax. Deprea se
caracteriza por poseer corolas infundibuliformes con lbulos de
menor longitud que el tubo y flamentos estaminales general-
mente ms largos que las anteras expandindose gradualmente
hacia su parte basal, a diferencia con Larnax que posee corolas
campanulado-rotadas con lbulos iguales o mayores que el tubo
y flamentos usualmente ms cortos o de igual longitud que las
anteras expandindose abruptamente hacia su base (Barboza &
Hunziker 1994, Sawyer 2007).
Los frutos de Deprea son bayas cubiertas por el cliz presisten-
te y acrescente una caracterstica tambin encontrada en Larnax
y otros gneros de la misma tribu (Hunziker 2001).
Deprea incluye actualmente ocho especies del noroeste de
Sudamrica (Leiva et al. 2005, Garzn-Venegas & Orozco 2007,
Sawyer 2007). Deprea bitteriana (Werderm.) Sawyer & Benitez
y D. orinocensis (Kunth) Raf. son las especies de distribucin
ms amplia, ya que se encuentran en Venezuela, Colombia y
Ecuador (Sawyer 1998). Deprea paneroi Benitez & M. Mart-
nez es endmica de Venezuela (Bentez de Rojas & Martnez
1992), mientras que D. nubicola Sawyer y D. cyanocarpa J.
Garzon & C. I. Orozco se conocen hasta el momento solo de
Colombia (Garzn-Venegas & Orozco 2007, Sawyer 2007).
Deprea ecuatoriana Hunz. & Barboza es endmica del Ecuador
(Barboza & Hunziker 1996, Montfar & Pitman 2004), y D.
cuyacensis (Sawyer & S. Leiva) S. Leiva & P. Lezama crece en
el departamento de Piura, al norte del Per (Leiva et al. 2005).
Finalmente, D. cardenasiana Hunz. muestra una distribucin
muy disyunta del resto de las especies, ya que es endmica de
Bolivia (Hunziker 1977).
La mayora de las especies habitan preferentemente en los
bosques nublados, desde los 1500 hasta los 3500 m de elevacin
(Barboza & Hunziker 1994). Son subarbustos o arbustos, con
fores de corolas infundibuliformes blanquecinas, verdosas, ama-
rillas, prpuras o violetas (Barboza & Hunziker 1994). Deprea
ecuatoriana es la nica especie del gnero con caractersticas
morfolgicas muy peculiares en cuanto a forma y color de sus
fores, ya que su corola es tubular-urceolada y anaranjada, lo que
la hace muy distinta del resto de las especies conocidas (Barboza
& Hunziker 1996). Segn Sawyer & Bentez de Rojas (1998) y
Sawyer (2005), D. ecuatoriana podra encontrarse tambin en
el norte del Per, cerca a la frontera con el Ecuador, pero hasta
la fecha no ha sido colectada en Per.
Teniendo en cuenta que el Per alberga un considerable
porcentaje de los bosques nublados de Sudamrica (Young &
Len 1999, 2000), resulta curioso el escaso nmero de especies
de Deprea en el territorio peruano, no existiendo hasta el mo-
mento informacin alguna sobre este gnero en los bosques de
la parte central y sur del pas.
En la parte central del Per, en el Departamento de Pasco, se
encuentran varias reas naturales de importancia para la conser-
vacin, entre ellas el Parque Nacional Yanachaga-Chemilln, que
debido a su accidentada topografa y variada vegetacin, donde
los Bosques nublados son la zona de vida ms representativa,
alberga una gran diversidad forstica an desconocida para la
ciencia, siendo las Solanceas una de las familias botnicas ms
diversas dentro de su fora (Vsquez et al. 2005).
Como parte de un estudio sobre la diversidad y distribucin
de la familia Solanaceae dentro del Parque Nacional Yanachaga-
144
Cueva &Trevio
Rev. peru. biol. 19(2): 143 - 147 (Agosto 2012)
Chemilln se pudo identifcar en el herbario HOXA un espci-
men de Deprea muy parecido a D. ecuatoriana. Sin embargo, al
revisar el material tipo de esta ltima especie result ser diferente
morfolgicamente, por lo que la describimos aqu como nueva
especie, informando adems sobre su distribucin y ecologa.
Descripcin taxonmica
Deprea oxapampensis M. Cueva & I. Trevio sp. nov.
(Figura 1)
TIPO: PER. Dpto. Pasco, Prov. Oxapampa, Distrito de
Huancabamba, Sector San Daniel, Parque Nacional Yanachaga-
Chemilln, parte alta de la trocha Erica, 3260 m, 1025'58"S
7526'11"W, 10 noviembre 2007, L. Hernani A., D. Rodrguez
M., A. Pea C.& R. Rivera C. 486 (holotipo: HOXA; Isotipos:
USM, HUT, MO).
Deprea ecuatoriana afnis, folia ellipticis glabrescentes, 3-6
foribus axillae, pedicelli foriferi elongatis et corolla 5 lobato difert.
Sufrtice ca. 0,5 m de alto. Tallos jvenes delgados, cilndri-
cos, de 12 mm en dimetro, ligeramente fexuosos, con entre-
nudos de 0,61,7 cm de longitud, verdosos o verde-purpreo
en las partes ms apicales, pubescentes, pelos blanquecinos
translcidos glandulares y no glandulares, los no glandulares
simples de 25 clulas uniseriadas, de 0,5 mm de longitud, los
glandulares de 0,8 mm de longitud de cabeza glandular y pie
multicelular uniseriado. Hojas simples, enteras, elpticas de
6,57,8 1,92,8 cm, de pice acuminado, base atenuada o
cortamente atenuada, en cada semilimbo con 78 venas secun-
darias distintas, al igual que la vena principal, superfcie adaxial
glabrescente, con pelos dispersos en la lmina pero ms densos en
la vena principal, pelos simples de 45 (6) clulas uniseriadas,
de 0,51 mm de longitud, superfcie abaxial con pubescencia
ms densa, pelos simples de 25 clulas uniseriadas, restringidos
mayormente a la vena principal, a las venas secundarias y a las
terciarias; pecolos de 310 mm de longitud. Inforescencia
axilar, fores en fascculos, 36 por axila; pedicelos en la antesis
de 1119 mm de longitud, 0,6 mm en dimetro en su parte
basal, 1 mm en dimetro en la parte apical, verdosos, purpreos
hacia el receptculo, pubescentes con pelos de 0,250,5 mm de
longitud, simples, pluricelulares y uniseriados, o glandulares de
pie pluricelular uniseriado y cabeza glandular; pedicelos en el
fruto de hasta 24 mm de longitud; botones jvenes con la corola
blanca que se torna amarillenta con la edad; receptculo atro-
purpreo. Flores con cliz cupuliforme, verdoso, de apariencia
truncada, con 5 lbulos cortos, triangulares, de pices obtusos,
de 0,5 mm de longitud y 0,5 mm de ancho, tubo de 2,2 mm
de longitud y 3,3 mm en dimetro, pubescente, con pelos sim-
ples uniseriados de hasta 5 clulas, glabro en su interior; corola
tuboloso-urceolada, anaranjada, de 9,5 mm de longitud, limbo
angosto pentalobado, lbulos triangulares muy cortos, de 1 mm
de longitud y 1,2 mm de ancho, superfcie externa de la corola
pubescente distalmente y glabrescente hacia la base del tubo,
interiormente glabra, pelos simples pluricelulares uniseriados de
0,250,4 mm de longitud; estambres insertados a 3 mm de la
base del tubo corolino sin ensanchamiento abrupto o apndice
alguno, flamentos estaminales glabros 4, iguales en longitud
1 ligeramente ms pequeo, parte libre del flamento de 2,5 mm
de longitud, de 2 mm de longitud en el estambre pequeo, de
0,7 mm en dimetro, anteras de 1,5 mm de longitud y 1 mm de
ancho, glabras, con el conectivo de hasta 0,5 mm de ancho, con
dehiscencia longitudinal; ovario de 2 mm de longitud y 1,31,4
mm en dimetro, glabro, estilo de 4,55 mm de longitud, 0,5
mm en dimetro, glabro, estigma de 1 mm de longitud. Fruto
baya elipsoidal de 6 mm de longitud y 4 mm en dimetro; en-
vuelto completamente por el cliz acrescente adpreso al fruto, de
7 mm de longitud y 4 mm en dimetro. Semillas no observadas.
Distribucin y ecologa
Hasta ahora slo se conoce de una colecta en la cordillera
Yanachaga, en el Parque Nacional Yanachaga-Chemilln. Crece
a 3260 m de altitud, en una zona de difcil acceso en bosque
nublado y en zonas de transicin de bosque esclerflo y praderas
expuestas o pajonales. En febrero del 2009 se realiz la bsqueda
de esta especie en distintos puntos del sector San Daniel durante
siete das (Figura 2 E), principalmente a lo largo de la denomi-
nada trocha Erica, lugar donde fue colectada esta especie por
primera vez, sin que se encontrara ningn nuevo individuo.
Por este motivo se le ha considerado como muy rara en la zona,
posiblemente restringida a las partes altas de la Cordillera, siendo
actualmente su estatus de conservacin incierto debido a la falta
de colecciones adicionales que puedan dar luz sobre el estado
real de su poblacin.
Etimologa
El epteto especfco alude a la provincia de Oxapampa,
que presenta una gran diversidad forstica, y privilegiada por
albergar cuatro reas naturales protegidas: el Parque Nacional
Yanachaga-Chemilln, la Reserva Comunal Yanesha, el Bosque
de Proteccin San Matas - San Carlos y parte de la Reserva
Comunal El Sira.
Discusin
Deprea oxapampensis es muy parecida a D. ecuatoriana, con la
que comparte corolas tubuloso-urceoladas de color anaranjado
y frutos elipsoidales con el cliz acrescente adpreso a la baya
(Figura 2 A-D). Sin embargo, se diferencia de esta ltima por
tener hojas elpticas de 6,57,8 cm de longitud y 1,92,8 cm de
ancho, 36 fores por axila, pedicelos forales en antesis de 1119
mm de longitud y corolas con 5 lbulos. Deprea ecuatoriana tiene
hojas ovadas de 2,23 cm de longitud y 1,41,6 cm de ancho,
2 3 fores axilares, pedicelos forales de 69 mm de longitud
y corolas con 5 lbulos que alternan con 5 lobulillos (Barboza
& Hunziker 1996). Otras caractersticas que las diferencian son
la coloracin de los pedicelos y del receptculo foral, as como
el tamao del cliz y la relacin de la longitud flamento/antera.
En D. ecuatoriana los pedicelos forales y el receptculo foral
son verdosos y el cliz es de 3,24 mm de longitud (Barboza
& Hunziker 1996), mientras que en D. oxapampensis poseen
una coloracin prpura y el cliz solo alcanza 2,7 mm como
mximo. As mismo, la longitud de la parte libre del flamento
de los estambres en D. ecuatoriana es 2,5 veces ms larga que
las anteras (Barboza & Hunziker 1996), mientras que en D.
oxapampensis es solo de 1,7. Otra peculiaridad observada en
D. oxapampensis es la diferencia de tamao de los flamentos
estaminales, donde cuatro de estos son de igual longitud y solo
uno es ligeramente pequeo. Como no se examinaron ms fores
de D. oxapampensis que las del tipo, es imposible saber el grado
de variacin de este carcter. En D. ecuatoriana los flamentos
son siempre de la misma longitud, que es lo ms habitual en el
gnero (Sawyer 2005).
145
Nueva especie de Deprea (Solanaceae) del Per
Rev. peru. biol. 19(2): 143 - 147 (August 2012)
Figura 1.Deprea oxapampensis M. Cueva & I. Trevio (L. Hernani et al. 486). A. Hbito; B. Flor en antesis; C. Fruto; D. Tricomas; E. Gineceo;
F. Anteras de izquierda a derecha en vista dorsal, lateral y ventral respectivamente; G. Flor abierta.
146
Cueva &Trevio
Rev. peru. biol. 19(2): 143 - 147 (Agosto 2012)
Figura 2. A,C. Deprea oxapampensis M. Cueva & I. Trevio (L. Hernani et al. 486; fotografa tomada por Antonio Pea). B, D. Deprea ecuato-
riana Hunz. & Barboza (fotografa tomada por P.M. Jrgensen de su colecta n 2204; disponible en http://www.tropicos.org/Specimen/1515691).
E. Vista de una parte de la cordillera Yanachaga en el sector San Daniel, habitat de D. Oxapampensis.
Figura 3. A. Tipo de Deprea oxapampensis M. Cueva & I. Trevio, L.Hernani et al. 486 (HOXA); B. Holotipo de Deprea ecuatoriana Hunz. &
Barboza, Len & Young 3379 (QCA).
147
Nueva especie de Deprea (Solanaceae) del Per
Rev. peru. biol. 19(2): 143 - 147 (August 2012)
Otro carcter interesante es que, al secarse, las fores de D.
oxapampensis tienden a perder la forma tubular-urceolada de la
corola, mientras que esto no ocurre en D. ecuatoriana, en la que
mantienen su forma aun despus del prensado (Figura 3 A, B).
Esta diferencia puede deberse a la existencia de lobulillos que
se alternan con los cinco lbulos principales de la corola en D.
ecuatoriana, que generan unos delgados pliegues en la superfcie
de la corola, lo que la hace ms fexible al prensado, a diferencia
de D. oxapampensis, en la cual las corolas tienden a desgarrarse
al ser presionados.
Caractersticas de D. oxapampensis son tambin su forma de
crecimiento y la disposicin de sus hojas en el tallo. El espcimen
examinado y las fotografas de las plantas en vivo muestran que
las hojas se disponen siempre en la parte apical de los tallos
delgados y ligeramente fexuosos. Este hbito recuerda a D.
paneroi, que se diferencia fcilmente por presentar una corola
prpura infundibuliforme notablemente pubescente en su inte-
rior (Bentez de Rojas 1992). Deprea nubicola comparte con D.
oxapampensis el tipo de fruto, con un cliz acrescente adpreso
a la baya, y la longitud de los pedicelos forales. Sin embargo,
D. nubicola tiene una corola prpura infundibuliforme de gran
tamao (1325 mm de longitud) y un fruto globoso, que la
diferencian de inmediato (Sawyer 2007).
El resto de especies de Deprea son fcilmente distinguibles
de D. oxapampensis. Deprea bitteriana, D. cardenasiana, D.
orinocensis, D. cuyacensis y D. cyanocarpa tienen corolas infundi-
buliformes con coloraciones que van de blanquecinas, amarillas,
prpuras o violetas, y frutos con un cliz acrescente que encierra
holgadamente a la baya (Garzn-Venegas & Orozco 2007, Leiva
et al. 2005, Sawyer 2007).
Aunque hasta el momento no se conocen las semillas de D.
oxapampensis, los caracteres mencionados distinguen claramente
a esta especie del resto de las del gnero y garantizan que nos
encontramos con un taxn que an no haba sido descrito.
La distribucin muy restringida y el escaso material colectado
hasta el momento de esta nueva especie es una situacin que
tambin se presenta en otras especies de la familia, en especial
de las que habitan en las zonas montaosas de los neotrpicos
donde el nivel de rareza o endemismo es alto (Knapp 2002).
Dentro del gnero igualmente existen casos de algunas especies
de las que se conoce poco sobre su distribucin y estado de
conservacin como D. nubicola y D. cardenasiana que fueron
descritas en base a una o dos colectas y hasta el momento solo se
conocen poblaciones restringidas a ciertas localidades de Colom-
bia y Bolivia respectivamente (Hunziker 1977, Sawyer 2007).
Al publicar esta especie en base a un solo espcimen se espera
estimular las colecciones en esta zona ya que la identifcacin y
descripcin de especies son los primeros pasos necesarios para
desarrollar objetivos e iniciar planes de conservacin a nivel
nacional y local (Mace 2004).
Agradecimientos
Expresamos nuestro agradecimiento al Dr. Jess Muoz
Fuente por la revisin de este trabajo; a la Blgo. Liz Hernani
Aldude, Blgo. Massiel Corrales Medina y a Antonio Pea Cruz
por proporcionarnos informacin y fotografas que ayudaron en
la descripcin de esta especie. A Rigoberto Rivera por el apoyo
en el trabajo de campo. A Katya Romoleroux, del herbario
QCA del Ecuador por permitirnos revisar el material tipo de D.
ecuatoriana. Al Ing. Rodolfo Vsquez y a la Blgo. Rocio Rojas
del Jardn Botnico del Missouri-Per y a todo el personal que
labora en esta institucin por su invalorable ayuda.
Literatura citada
Barboza G. E.& A. Hunziker. 1994. Estudios sobre Solanaceae
XXXVII. Sinopsis taxonmica de Deprea. Kurtziana 23:
101-124.
Barboza G. E.& A. Hunziker. 1996. Estudio sobre Solanaceae XLII.
Una nueva especie de Deprea del Ecuador. Darwiniana 34
(1-4): 109-111.
Bentez de Rojas C. 1992. A new species of Deprea (Solanaceae)
from Venezuela. Phytologia 73 (4): 270-273.
Garzn-Venegas J. & C. I. Orozco. 2007. Deprea cyanocarpa (So-
lanaceae: Physaleae): una nueva especie para Colombia.
Caldasia 29(2): 219-227.
Hunziker A. 1977. Estudio sobre Solanaceae. VIII. Novedades va-
rias sobre Tribus, Gneros, Secciones y Especies de Sud
Amrica. Kurtziana 10: 7-50.
Hunziker A. 2001. Genera Solanacearum. The genera of Solanaceae
ilustrated, arranged according to a new system.A.R.G.
Gantner Verlag K.-G.
Knapp S. 2002. Assessing Patterns of Plant Endemism in Neotropical
Uplands. The Botanical Review 68(1):22-37.
Leiva S., P. Lezama A. & M. Zapata. Primera especie de Deprea
Rafnesque (Solanaceae: Solaneae) en Per. Arnaldoa 12
(1-2): 62-66.
Mace G. 2004. The role of taxonomy in species conservation. Phil.
Trans. R. Soc. Lond. B 359:711-719.
Montfar R.& Pitman, N. 2004.Deprea ecuatoriana.En: IUCN 2011.
IUCN Red List of Threatened Species.Version 2011.2.
<www.iucnredlist.org>. visitado en 16 April 2012.
Sawyer N. W. 2001.New species and combinations in Larnax (So-
lanaceae).Novon 11: 460-471.
Sawyer N. W. 2005. Systematics of Deprea and Larnax(Solanaceae).
En: R. C. Keating, C. V. Hollowell& T. B. Croat (eds.),
Festschrift for William G. DArcy: The Legacy ofa taxono-
mist. Monographs in Systematic Botany from the Missouri
Botanical Garden 104: 259-285.
Sawyer N. W. 2007. Deprea nubicola (Solanaceae), a new species
from northern Colombia. Brittonia 59 (1): 54-56.
Sawyer N. W. & C. Bentez de Rojas. 1998. Morphological analysis
of three equivocal sibling species of Deprea(Solanaceae).
Brittonia 50 (4): 524-535.
Vsquez R., R. Rojas, A. Monteagudo, et al. 2005. Flora vascular
de la Selva Central del Per: una aproximacin de la
composicin forstica de tres reas Naturales Protegidas.
Arnaldoa 12 (1-2): 112-125.
Young K. R. & B. Len. 1999. Perus humid eastern montane forest:
An overview of their physical settings, biological diversity,
human use and settlement, and conservation needs. DIVA
Technical Report 5: 1-97.
Young, K. R.& B. Len.2000. Biodiversity Conservation in Perus
Eastern Montane Forests. Mountain Research and Deve-
lopment 20 (3): 208-211.
148
Cueva &Trevio
Rev. peru. biol. 19(2): 143 - 147 (Agosto 2012)
149
Flora vascular de las lomas de Villa Mara y Amancaes
Rev. peru. biol. 19(2): 149 - 158 (August 2012)
Rev. peru. biol. 19(2): 149 - 158 (Agosto 2012)
Facultad de Ciencias Biolgicas UNMSM
ISSN 1561-0837
Flora vascular de las lomas de Villa Mara y Amancaes, Lima, Per
Huber Trinidad
1
, Elluz Huamn-Melo
1
, Amalia Delgado
1
y Asuncin Cano
1,2
Vascular fora from Villa Mara and Amancaes Lomas, Lima, Peru
1 Laboratorio de Florstica, Depar-
tamento de Dicotiledneas, Museo
de Historia Natural Universidad
Nacional Mayor de San Marcos.
Av. Arenales 1256, Lima 11, Per.
Email Huber Trinidad Patricio:
htrinidadpatricio@gmail.com
Email Asuncin Cano:
ashuco@yahoo.com
2 Instituto de Investigacin de Cien-
cias Biolgicas Antonio Raimondi
(ICBAR), Facultad de Ciencias
Biolgicas, Universidad Nacional
Mayor de San Marcos.
Presentado: 22/02/2012
Aceptado: 28/09/2012
Publicado online: 10/11/2012
Resumen
En el presente trabajo se informa sobre el inventario de la fora vascular de las lomas de Villa Mara y Aman-
caes, Lima. Las expediciones para inventariar ambas localidades se realizaron en setiembre y octubre del
2011. En ambos lugares fueron registradas 121 especies, agrupadas en 41 familias y 99 gneros. Las
familias ms diversas fueron Asteraceae y Boraginaceae con 17 y 8 especies respectivamente. En las
lomas de Villa Mara se registraron 112 especies agrupadas en 99 gneros y 39 familias, de las cuales 20
fueron endmicas; siendo una de las lomas ms diversas de Lima despus de Lachay y Carabayllo. En
las lomas de Amancaes se registraron 51 especies agrupadas en 43 gneros y 28 familias, de las cuales
9 fueron endmicas. El bajo nmero de especies en Amancaes se debe a la reduccin del rea que ocupa
producto del crecimiento urbano. En ambas localidades, la vegetacin es predominantemente herbcea, con
el 77% de las especies en Villa Mara y 86% en Amancaes. Ambas lomas estn sometidas un gran impacto
por la expansin urbana, lo que ha reducido el tamao de sus reas y ha contribuido a su contaminacin,
principalmente con los residuos slidos.
Palabras clave: Lomas, Villa Mara, Amancaes, Conservacin, Endemismos.
Abstract
This paper reports the inventory of vascular fora of Villa Maria and Amancaes Lomas, Lima, Peru. The expedi-
tions for inventorying both locations were conducted in September and October 2011. We registered 121 spe-
cies in both localities, belonging to 99 genera and 41 families. The most diverse families were Asteraceae and
Boraginaceae with 17 and 8 species respectively. Villa Maria lomas 112 species in 99 genera and 39 families
were registered, of which 20 are endemic. Villa Maria lomas is one of the most diverse lomas in Lima after
Lachay and Carabayllo. In Amancaes lomas, 51 species in 43 genera and 28 families were registered, of which
9 are endemic. The low number of species in Amancaes lomas is due to the reduction of the area occupied,
caused by urban growth. In both localities, the vegetation is predominantly herbaceous, with 77% of the species
in Villa Maria and 86% in Amancaes. Both lomas are having a high impact by the urban expansion, which has
reduced their area size and contributed to their pollution, mainly with solid waste.
Keywords: Lomas, Villa Mara, Amancaes, Conservation, Endemic.
Introduccin
Las lomas son formaciones vegetales nicas que se presentan
desde la costa norte del Per (8S) hasta el norte de Chile (30S)
(Ferreyra 1993, Rundell et al. 1991). Se desarrollan en laderas
y quebradas pedregosas orientadas hacia el mar, donde la
principal fuente de humedad proviene de la condensacin
de neblina originada en el Ocano Pacfco (Ferreyra 1993,
Talavera et al. 2001). Se presentan como islas de vegetacin
separadas por distancias variables de hbitat completamente
rido, principalmente entre los 600 y 900 msnm (Ferreyra
1993, Dillon et al. 2003).
La fora vascular de las lomas es diversa y est caracterizada
por poseer un alto nmero de especies endmicas (Rundell
1991), siendo las familias Asteraceae, Fabaceae, Malvaceae,
Solanaceae y Poaceae, las ms diversas (Pfaur 1982). La
vegetacin est caracterizada por la abundancia de especies
herbceas anuales que tapizan totalmente la superfcie del
suelo durante los meses de mayor humedad. Adems, en la
mayora de las lomas, existe un estrato arbustivo distribuido
en forma dispersa o en algunos lugares ms densos y en los
sectores donde existe mayor captacin de neblina, se pueden
desarrollar especies arbreas (INRENA 1996).
La infuencia del hombre sobre las lomas ha sido signif-
cativa a lo largo del tiempo. Desde la antigedad estos eco-
sistemas tuvieron gran importancia como zonas de pastoreo.
Con respecto a lomas de los alrededores de la ciudad de Lima,
la principal amenaza es la expansin urbana, la misma que
viene desarrollndose de manera desordenada, afectando su
extensin; cobertura vegetal y diversidad forstica. El presente
trabajo informa sobre la composicin de la fora vascular de las
lomas de Villa Mara y Amancaes impactadas por el crecimiento
urbano de Lima.
rea de estudio
El presente estudio se realiz en las lomas de Villa Mara y
Amancaes. Las lomas de Villa Mara se encuentran ubicadas al
sureste de la ciudad de Lima, en el distrito de Villa Mara del
Triunfo (UTM 18L 290876 8657580), presentan altitudes
entre los 400 y 1000 m y junto con las lomas del Lcumo,
forman parte del antiguo sistema de lomas de Atocongo. Las
lomas de Amancaes se encuentran ubicadas al norte de la ciudad
de Lima, en el lmite de los distritos de Independencia, Rmac y
San Juan de Lurigancho (18L 278898 8672619) y presentan
altitudes entre 300 y 800 m (Fig. 1).
Materiales y mtodos
En setiembre y octubre del 2011, se realizaron evaluaciones
en cada una de las localidades; meses donde el desarrollo de la
vegetacin de lomas en la costa central es ptimo. Se realizaron
recorridos en todo el rea de ambas lomas estudiadas; regis-
trando, recolectando y fotografando todas las especies y las
comunidades vegetales. Los especmenes fueron recolectados
segn tcnicas estndares (Bridson & Forman, 1992); prensados
y trasladados al Museo de Historia Natural de la Universidad
150
Trinidad et al.
Rev. peru. biol. 19(2): 149 - 158 (Agosto 2012)
Nacional Mayor de San Marcos (MHN-UNMSM), para su
secado, determinacin y deposito en el Laboratorio de Florstica
del MHN-UNMSM.
Para las determinaciones, se utilizaron claves taxonmicas
como las de Macbride et al. (1936 y siguientes), Sagstegui y
Leiva (1993), Tovar (1993), entre otros; estas fueron corrobo-
radas, cuando fue necesario, por consulta a especialistas y/o
a las colecciones del Herbario San Marcos (USM) y bases de
datos de herbarios virtuales como las del Missouri Botanical
Garden (MO) y Field Museum Herbarium (F). El ordenamiento
taxonmico de las especies, se realiz utilizando el sistema de
clasifcacin de APGIII (Bremer et al. 2009).
Resultados
Diversidad forstica
Se registraron 121 especies de plantas vasculares en ambas lo-
mas, agrupadas en 99 gneros y 41 familias botnicas (Apndice
1), de las cuales 94 (77,7%) son hierbas, 14 (11,6%) arbustos,
cinco (4,1%) hierbas trepadoras, cuatro (3,3%) rboles y cuatro
(3,3%) suculentas (Tabla 1). Adiantum poiretii fue el nico
pteridfto registrado.
Las Magnolipsida son el grupo dominante con 107 especies
(88,4%), en 86 gneros (86,9%) y 33 familias (80,5%). Las
Liliopsida o Monocotiledneas estuvieron representadas con
13 especies (12,1%) en 12 gneros (12,1%) y siete familias
(17,1%). Las Asteraceae son las ms diversas con 17 especies en 16
gneros, seguidas por las Boraginaceae (8/3), Caryophyllaceae (6/6),
Euphorbiaceae (6/5), Fabaceae (6/6) y Solanaceae (6/5) (Fig. 2).
Diversidad forstica en lomas de Villa Mara.- Se regis-
traron 112 especies, agrupadas en 94 gneros y 39 familias.
Apndice 1 y Tabla 2. Se registr slo un pteridofto: Adiantum
poiretii; las 111 especies restantes pertenecen a la divisin Mag-
noliophyta, donde el 91% (100 especies) corresponden a la
clase Magnoliopsida y el 9% (11 especies) a la clase Liliopsida
(Apndice 1). Las familias ms diversas fueron Asteraceae con
17 especies en 16 gneros, Boraginaceae (8/3), Euphorbiaceae
(6/5), Fabaceae (6/6) y Solanaceae (6/5) (Fig. 3). Los gneros con
mayor nmero de especies fueron Heliotropium (Boraginaceae)
y Erodium (Malvaceae) con cinco (4,4%) y tres (2,7%) especies
respectivamente, los otros gneros solo presentaron una o dos
especies. Con respecto a las forma de crecimiento, 86 especies
fueron hierbas, 13 arbustos, seis hierbas trepadoras, cuatro
rboles y tres suculentas.
Diversidad forstica en lomas de Amancaes.- Se registraron
51 especies de plantas vasculares, agrupadas en 43 gneros y 28
familias. Apndice 1 y Tabla 2. Las Magnolipsidas represen-
taron la vegetacin dominante, albergando el 94% del total de
las especies. Las familias ms diversas fueron Asteraceae con 9
especies en 9 gneros, Solanaceae (4/3), Cactaceae (3/2) y Car-
yophyllaceae (3/3) (Fig. 4). Los gneros con mayor nmero de
especies fueron: Chenopodium (Amaranthaceae), Cleistocactus
(Cactaceae), Erodium (Geraniaceae), Fuertesimalva (Malvaceae),
Oxalis (Oxalidaceae), Solanum (Solanaceae) y Salvia (Lamiaceae)
con dos especies cada uno; los gneros restantes registraron
solo una especie. Con respecto a las forma de crecimiento, 44
especies fueron hierbas, tres suculentas, dos arbustos, una hierba
trepadora y un rbol.
Forma de crecimiento N especies %
Hierba (H) 94 77,69
Arbusto (Ab) 13 10,74
Trepador (Tr) 6 4,96
Suculenta (Su) 4 3,31
rbol (Ar) 4 3,31
Tabla 1. Forma de crecimiento de las especies registradas en las
lomas de Amancaes y Villa Mara, septiembre-octubre 2011
17
8
6 6 6 6
5 5
62
16
3
6
5
5
6
4
3
51
0
10
20
30
40
50
60
70
A
s
t
e
r
a
c
e
a
e
B
o
r
a
g
i
n
a
c
e
a
e
F
a
b
a
c
e
a
e
E
u
p
h
o
r
b
i
a
c
e
a
e
S
o
l
a
n
a
c
e
a
e
C
a
r
y
o
p
h
y
l
l
a
c
e
a
e
L
a
m
i
a
c
e
a
e
A
m
a
r
a
n
t
h
a
c
e
a
e
O
t
r
o
s
N
.
E
s
p
e
c
i
e
s
y
g
n
e
r
o
s
Especies
Gneros
Figura 2. Familias con mayor nmero de especies y gneros en las
lomas de Villa Mara y Amancaes, setiembre-octubre 2011.
Villa Mara Amancaes
Familias 39 29
Gneros 94 44
Especies 112 51
Tabla 2. Nmero de Familias gneros y especies registradas en las
lomas de Amancaes y Villa Mara, septiembre-octubre 2011
Figura 1. Ubicacin de las lomas de Amancaes y Villa Mara.
Lomas de Villa Mara
Lomas de Amancaes
Callao
5 km
Ocano Pacfico
Lima
Metropolitana
151
Flora vascular de las lomas de Villa Mara y Amancaes
Rev. peru. biol. 19(2): 149 - 158 (August 2012)
Endemismos
En las lomas de Villa Mara, se registraron 20 especies end-
micas para el Per, las cuales representan el 17,8% del total de
especies registradas para esta loma; donde la familia Cactaceae
registr cuatro especies, Asteraceae, Solanaceae y Euphorbia-
ceae, tres cada una. En las lomas de Amancaes, se registraron 9
taxones endmicos, que representan el 17,6% del total de las
especies registradas para esta loma, donde la familia Cactaceae
fue la mejor representada con tres especies. Stenomesson favum
(Amaryllidaceae) fue la nica Liliopsida endmica. Tabla 3.
Notas sobre la vegetacin
Lomas de Villa Mara.- Hacia el lado occidental de las
lomas, en donde las masas de niebla inciden directamente, la
humedad es mayor, favoreciendo un mejor desarrollo de la
vegetacin, registrndose extensas poblaciones de Nasa urens
(Loasaceae) y Fuertesimalva spp. (Malvaceae). En la parte
baja de la ladera se pueden encontrar extensas poblaciones
de Sicyos baderoa (Cucurbitaceae), una especie voluble que se
encuentra alternando con poblaciones de Fuertesimalva spp.,
Salvia spp. (Lamiaceae) y Nicotiana paniculata (Solanaceae),
17
8
6 6 6
5 5 5
54
16
3
5
6
5
3
4
5
47
0
10
20
30
40
50
60
A
s
t
e
r
a
c
e
a
e
B
o
r
a
g
i
n
a
c
e
a
e
S
o
l
a
n
a
c
e
a
e
F
a
b
a
c
e
a
e
E
u
p
h
o
r
b
i
a
c
e
a
e
A
m
a
r
a
n
t
h
a
c
e
a
e
L
a
m
i
a
c
e
a
e
C
a
r
y
o
p
h
y
l
l
a
c
e
a
e
O
t
r
o
s
N
.
E
s
p
e
c
i
e
s
y
g
n
e
r
o
s
Especies
Gneros
17
8
6 6 6
5 5 5
54
16
3
5
6
5
3
4
5
47
0
10
20
30
40
50
60
A
s
t
e
r
a
c
e
a
e
B
o
r
a
g
i
n
a
c
e
a
e
S
o
l
a
n
a
c
e
a
e
F
a
b
a
c
e
a
e
E
u
p
h
o
r
b
i
a
c
e
a
e
A
m
a
r
a
n
t
h
a
c
e
a
e
L
a
m
i
a
c
e
a
e
C
a
r
y
o
p
h
y
l
l
a
c
e
a
e
O
t
r
o
s
N
.
E
s
p
e
c
i
e
s
y
g
n
e
r
o
s
Especies
Gneros
Figura 3. Familias con mayor nmero de especies presentes en las
lomas de Amancaes, setiembre-octubre 2011.
Figura 4. Familias con mayor nmero de especies presentes en las
lomas de Villa Mara, setiembre-octubre 2011.
Familia Especie Villa Mara Amancaes
Amaryllidaceae Stenomesson favum Herb. X
Amaranthaceae Atriplex rotundifolia Dombey ex Moq. X
Asteraceae Ophryosporus pubescens (Sm.) R.M. King & H. Rob. X
Philoglossa peruviana DC. X X
Senecio abadianus DC. X
Boraginaceae Cryptantha limensis I.M. Johnst. X
Heliotropium pilosum Ruiz & Pav. X
Cactaceae Cleistocactus acanthurus (Vaupel) D.R. Hunt X X
Cleistocactus sextonianus (Backeb.) D.R. Hunt X
Haageocereus limensis (Salm-Dyck) F. Ritter X X
Haageocereus multangularis (Haw.) F. Ritter X
Calceolariaceae Calceolaria angustifora Ruiz & Pav. X
Caprifoliaceae Valeriana pinnatifda Ruiz & Pav. X
Caryophyllaceae Drymaria paposana var. weberbaueri (Muschl.) Duke X
Euphorbiaceae Andrachne microphylla (Lam.) Baill. X
Cistanthe lingulata (Ruiz & Pav.) Hershkovitz X
Cistanthe paniculata (Ruiz & Pav.) Carolin ex Hershkovitz X
Loasaceae Mentzelia scabra subsp. grandifora (Ruiz & Pav. ex G. Don) Weigend X
Montiaceae Calandrinia alba (Ruiz & Pav.) DC. X X
Oxalidaceae Oxalis lomana Diels X
Solanaceae Exodeconus prostratus (L'Hr.) Raf. X
Nicotiana paniculata L. X X
Nolana humifusa (Gouan) I.M. Johnst. X X
Tabla 3. Lista de especies endmicas registradas en las lomas de Villa Mara y Amancaes, septiembre-octubre 2011, segn Len et al. 2006.
152
Trinidad et al.
Rev. peru. biol. 19(2): 149 - 158 (Agosto 2012)
adems se registraron algunos individuos arbreos de la fa-
milia Fabaceae, como Caesalpinia spinosa (Tara) y Parkinsonia
aculeata (Azote de Cristo). Algunas hierbas frecuentes fueron:
Raimundochloa trachyantha (Poaceae), Veronica persica (Plan-
taginaceae), Astrephia chaerophylloides (Caprifoliaceae), Cal-
ceolaria pinnata (Calceolariaceae), Stellaria media, Cerastium
glomeratum (Caryophyllaceae) y Emex spinosa (Polygonaceae),
esta ltima es una especie invasora extica fcilmente reco-
nocible por sus frutos trgonos espinescentes. Protegidas por
las rocas; asimismo, es comn encontrar a Parietaria debilis
(Urticaceae), y escasamente Adiantum poiretii (Pteridaceae),
el nico helecho registrado.
En la parte media, empiezan a hacerse ms evidentes extensas
poblaciones de Nasa urens que adornan toda la loma con sus
llamativas fores amarillas; otras especies bastante abundantes
son Solanum montanum, tambin existen aqu poblaciones
grandes de Fuertesimalva spp. Entre otras especies frecuentes
encontramos Heliotropium arborescens, Cyclanthera mathewsii
(Cucurbitaceae), Sysimbrium orientale (Brassicaceae), Erigeron
leptorhizon (Asteraceae), Solanum peruvianum, Sonchus oleraceus,
Acmella oleracea, Phyloglossa peruviana (Asteraceae) y Raimun-
dochloa trachyantha.
En las zonas rocosas y quebradas pedregosas se registr a
Begonia octopetala (Begoniaceae), adems de cuatro individuos
de Vasconcellea candicans Mito (Caricaceae) una especie
arborescente caracterizada por presentar un fruto comestible
consumido comnmente por los pobladores locales.
Conforme se asciende, se empiezan a observar individuos del
arbusto Croton alnifolius (Euphorbiaceae), hacindose bastante
frecuente en la parte alta de la loma, formando matorrales
laxos en algunas partes. Algunas especies bastante frecuentes
fueron Alternanthera pubifora (Amaranthaceae), Nolana humi-
fusa, Solanum peruvianum (Solanaceae) y Haageocereus limensis
(Cactaceae).
Hacia el lado oriental de las lomas, la humedad es menor
y la vegetacin es rala, algunas veces restringida a quebradas
pedregosas. Se pueden observar algunas cactceas columnares
como Haageocereus limensis, H. multangularis y Cleistocactus
acanthurus, y algunas bromeliceas tales como Puya ferruginea
y Tillandsia latifolia; todas estas especies son xerofticas. En las
partes altas existen matorrales de Croton alnifolius que se encuen-
tra alternando algunas veces con Haaggeocereus limensis; otras
especies frecuentes son: Crotalaria incana (Fabaceae), Baccharis
salicifolia (Asteraceae), Jacquemontia unilateralis (Convolvula-
ceae), Eragrostis attenuata (Poaceae), Cryptantha parvifora y C.
limensis (Boraginaceae). En las quebradas se pueden encontrar
frecuentemente especies como Heliotropium rufpilum, Loasa
ntida y Cyclanthera sp. y de manera escasa Ipomoea dumetorum
(Convolvulaceae), Alstroemeria lineatifora (Alstroemeriaceae),
entre otras.
Lomas de Amancaes.- La vegetacin de la ladera occidental
(hacia el distrito del Rmac e Independencia), en su parte inferior,
est conformada predominantemente por Sicyos baderoa, la cual
crece intercalada con parches de Nasa urens, Fuertesimalva spp.,
Erodium spp. y Salvia spp. A medida que se va ascendiendo, entre
las rocas se pueden apreciar pequeas poblaciones de Begonia
octopetala. Asimismo se pueden distinguir parches de Calceola-
ria pinnata y Fumaria capreolata (Papaveraceae) alternadas con
Philoglossa peruviana (Asteraceae).
En las partes ms altas, con menor incidencia de neblina, la
cobertura de vegetacin es menor, siendo las especies ms fre-
cuentes Nicotiana paniculata, Nasa urens, Solanum montanum,
Nolana humifusa, Oxalis lomana y Loasa nitida.
En las laderas orientadas hacia el distrito de San Juan de
Lurigancho, debido a las condiciones ambientales ms xerftas,
la cobertura vegetal es menor en comparacin con la ladera occi-
dental. En la parte ms baja de la loma, la especie ms frecuente
fue Nicotiana paniculata, creciendo intercalada con parches de
Nasa urens. Hacia las partes altas, se han registrado Tillandsia
latifolia, Cleistocactus acanthurus, Cleistocactus sextonianus, Ha-
ageocereus limensis creciendo entre rocas.
Estado de Conservacin
En las lomas de Villa Mara, segn el DS 043-AG (2006),
se han registrado ocho especies bajo algn grado de amenaza,
destacndose la presencia de Vasconcellea candicans, una especie
que se encuentra en peligro crtico (CR), de la cual en el presente
trabajo slo se han registrado cuatro individuos.
En la loma de Amancaes, se han registrado tres especies bajo
algn grado de amenaza; una en situacin vulnerable (VU) y
dos en peligro (EN). Anteriormente en esta loma se registraron
tambin algunos individuos (ocho) de Vasconcellea candicans
(Cuya y Snchez 1991), algunos de ellos fueron observados
por uno de los autores (AC) en el 2001, pero en este estudio no
fueron observados (Tabla 4). Es importante hacer notar que en
esta loma ya se ha documentado la extincin local de varias espe-
cies, entre ellas la especie simblica Ismene amancaes Amancay
(Amaryllidaceae). Tambin Len et al. (2002) documentaron
para esta loma cuatro especies de helechos que en la presente
evaluacin no fueron encontradas.
Familia Especies Categora Villa Mara Amancaes
Cactaceae Cleistocactus acanthurus(Vaupel) D.R. Hunt EN X
Cactaceae Cleistocactus acanthurus(Vaupel) D.R. Hunt EN X
Cactaceae Haageocereus limensis (Salm-Dyck) F. Ritter EN X X
Cactaceae Haageocereus multangularis (Haw.) F. Ritter EN X
Caricaceae Vasconcellea candicans (A. Gray) A. DC. CR X
Euphorbiaceae Cnidoscolus basiacanthus (Pax & K. Hoffm.) J.F. Macbr. VU X
Fabaceae Caesalpinia spinosa (Molina) Kuntze VU X
Fabaceae Caesalpinia spinosa (Molina) Kuntze VU X
Fabaceae Vachellia macracantha (Humb. & Bonpl. ex Willd.) Seigler & Ebinger NT X
Iridaceae Tigridia pavonia (L. f.) DC. NT X
Tabla 4. Lista de especies amenazadas en las lomas de Villa Mara y Amancaes segn el DS 043-2006
153
Flora vascular de las lomas de Villa Mara y Amancaes
Rev. peru. biol. 19(2): 149 - 158 (August 2012)
Impactos
Ambas localidades, por su cercana a la ciudad de Lima,
presentan un fuerte impacto antropognico producido por la
poblacin establecida en sus alrededores. Entre los impactos
ms importantes que podemos mencionar, la reduccin de sus
reas debido a la construccin de viviendas y vas de acceso, estas
ltimas favoreceran en el mediano o largo plazo su ocupacin de
las reas de lomas. En las lomas de Villa Mara, la construccin
de una carretera de ingreso hacia el extremo norte, est alterando
y fragmentando el paisaje, poniendo en peligro la cobertura
vegetal e incrementando el riesgo de una potencial invasin de
estos terrenos. Asimismo se observan antenas y restos de atra-
panieblas en un sector de las mismas, los cuales evidentemente
han sido abandonados despus de la ejecucin de su propsito.
Otros efectos de las actividades humanas, se ven refejados
en la presencia de algunas especies introducidas en las lomas.
Como en el caso de Villa Mara, donde se registr la presencia
de Tropaeolum majus (Tropaelolaceae), una planta, con variados
usos (ornamental, medicinal y alimenticio), que al encontrar
condiciones ptimas para su desarrollo, viene expandiendo su
distribucin a lo largo de la loma. Otras especies introducidas en
las lomas son: Casuarina equisetifolia (Casuarinaceae), Aptenia
cordifolia (Aizoaceae) y Arundo donax (Poaceae).
Discusin
El presente estudio es el primero realizado sobre diversidad
forstica presente en las lomas de Villa Mara, la misma que
por su elevado nmero de especies (112), podran considerarse
como una de las lomas con mayor diversidad en Lima, junto
con las lomas de Lachay con 144 especies (Cano et al. 1999) y
Carabayllo con 124 (datos no publicados).
En el nico inventario forstico publicado para las lomas de
Amancaes, Cuya y Snchez (1991) reportaron la presencia de 24
especies; de las cuales, en el presente estudio, no se han registrado
Tourretia lappaceae (Bignoniaceae) y Piqueria peruviana (Astera-
ceae); estas especies, posiblemente no se desarrollaron durante
el periodo de estudio al no encontrar condiciones ambientales
ptimas o pudieron haberse extinguido en el rea debido a la
continua disminucin en la extensin de las lomas. El nmero
de especies registradas es bajo (50 spp.) en comparacin con los
presentes en las lomas de Villa Mara o en otras lomas de Lima,
esto podra atribuirse a que actualmente es una de las lomas
de menor rea en el departamento, producto de la continua
expansin urbana que invade sus reas. Sin embargo, el registro
histrico de la composicin de la fora vascular de las lomas de
Amancaes superar las 100 especies (Cano et al. en preparacin),
confrmndose la extincin local de varias especies.
En este estudio se comprueba la importancia de la lomas
como ecosistemas nicos que albergan un nmero signifcativo
de especies endmicas. En ambas lomas se encontr que las es-
pecies endmicas signifcaban un porcentaje considerable de la
vegetacin presente (18% en cada loma), entre ellas cabe resaltar
a Senecio abadianus (Asteraceae), especie registrada nicamente
para el departamento de Lima.
En ambas localidades, la vegetacin es predominantemente
herbcea, el 77% de especies en las lomas de Villa Mara y 86%
en las lomas de Amancaes. Los arbustos son ms frecuentes
en las lomas de Villa Mara (14 spp.) en comparacin a las de
Amancaes (2 spp.), lo cual nos indicara que esta loma se en-
cuentra menos impactada, adems de poseer micro hbitats ms
variados por la mayor rea que posee; esto reafrmado tambin
por la presencia de especies arbreas; cuatro en las lomas de Villa
Mara (Vasconcellea candicans, Vachellia macracantha, Caesalpinia
spinosa y Parkinsonia aculeata) y una en las lomas de Amancaes
(Caesalpinia spinosa).
Debido a la cercana de las lomas al cinturn urbano, a las
fuertes presiones antrpicas que impactan sobre su extensin e
integridad, por albergar especies endmicas y protegidas por la
legislacin nacional es de urgente necesidad establecer planes
de manejo, programas de conservacin y polticas que permitan
conservar y proteger estos ecosistemas nicos del desierto costero
peruano-chileno.
Agradecimientos
Agradecemos a la ONG A Rocha Per, por el fnanciamiento
que hizo posible la realizacin de este estudio, adems por su
inters en fomentar la conservacin de este tipo de ecosistemas.
Un agradecimiento especial a Eduardo Navarro, Warner Aparco,
Anglica Rodrguez, Raquel Gonzles, Sabby Araujo y Edwin
Trinidad por su apoyo durante el trabajo de campo. Tambin
expresamos nuestra gratitud a Mnica Arakaki por su ayuda con
la revisin del abstract.
Literatura citada
Bremer B., K. Bremer, M. W. Chase, M. F. Fay, J. Reveal, D. E.
Soltis, P. S. Soltis & P. F. Stevens. 2009. An Update of the
Angiosperm Phylogeny Group Classifcations for Orders
and Families of Flowering Plants: APG III. Botanical
Journal of The Linneaus Society 161. Pp. 105-121.
Bridson D. & L. Forman. 1992 (eds.). The Herbarium Handbook.
Royal Botanic Gardens. Kew.
Cano A., J. Roque, M. Arakaki, C. Arana, M. I. La Torres, N. Llerena
& N. Refulio. 1999. Diversidad Florstica de las lomas
de Lachay (Lima) durante el evento "El Nio 1997-98".
En: El Nio 1997-98 y su impacto sobre los ecosistemas
Marino y Terrestre. J. Tarazona y E. Castillo (Eds). Rev.
peru. biol. Vol Extraordinario: 125-132 (1999)
Cuya O. & S. Snchez. 1991. lomas de Amancaes: lomas que deben
conservarse. Boletn de Lima, N76 pp 59-64.
Dillon M., Nakawaka M. & Leiva, S. 2003. The lomas formations of
Coastal Per: Composition and Biogeographic History. "
El Nio in Peru: Biology and Culture Over 10,000 Years".
Fieldiana: Botany. Chicago.43:1-9.
Ferreyra R. 1993. Registros de la vegetacin en la costa peruana en
relacin con el fenmeno el Nio. Boletn del Instituto
Francs de Estudios Andinos. 22 (1): 259-266. Argentina.
INRENA. 1996. Gua explicativa del mapa forestal. Direccin ge-
neral forestal. Ministerio de Agricultura. INRENA. Lima.
Len B., J. Roque, C. Ulloa-Ulloa, N. Pitman, P.M. Jorgensen & A.
Cano (editores). 2006. El libro rojo de las plantas peruanas
del Per. Rev. peru. biol. Nmero especial 13(2). Pg. 971.
Len B., A. Cano & K. R. Young. 2002. Los helechos de las lomas
costeras del Per. Arnaldoa 9(2):7-42.
Macbride J. Francis. et al. Flora of Peru. Botanical Series. Field
Museum of Natural History. Botanical Series. Chicago.
1936 y siguientes.
Pfaur J. 1982. Dynamycs of plant comunities in the lomas of
southern Per. Vegetation 49: 193-171. Netherland.
Rundell P., M. Dillon, B. Palma, H. Money, S. Gulmon, & J. Ehle-
ringer. 1991. The Phytogeography and ecology of the
coastal Atacama and Peruvian deserts. Aliso 13 (1): 1-50.
Sagstegui A. & Leiva, S. 1993. Flora invasora de los cultivos del
Per. 1nd ed. Trujillo, Editorial Libertad EIRL. Pp. 539.
154
Trinidad et al.
Rev. peru. biol. 19(2): 149 - 158 (Agosto 2012)
Taxa superior Especie Villa Mara Amancaes
Forma de
crecimiento
Clase: Liliopsida
Alstroemeriaceae
Alstroemeria lineatifora Ruiz & Pav. X H
Amaryllidaceae
Stenomesson coccineum Herb. X H
Stenomesson favum Herb. X H
Asparagaceae
Anthericum eccremorrhizum Ruiz & Pav. X H
Oziroe bifora (Ruiz & Pav.) Speta X H
Bromeliaceae
Puya ferruginea (Ruiz & Pav.) L.B. Sm. X H
Tillandsia latifolia Meyen X X H
Commelinaceae
Commelina fasciculata Ruiz & Pav. X H
Iridaceae
Tigridia pavonia (L. f.) DC. X H
Poaceae
Eragrostis attenuata Hitchc. X H
Lamarckia aurea (L.) Moench X H
Poa annua L. X H
Raimundochloa trachyantha (Phil.) A.M. Molina X X H
Clase: Magnoliopsida
Acanthaceae
Dicliptera montana Lindau X H
Amaranthaceae
Alternanthera halimifolia (Lam.) Standl. ex Pittier X H
Alternanthera pubifora (Benth.) Kuntze X H
Atriplex rotundifolia Dombey ex Moq. X H
Chenopodium murale L. X X H
Chenopodium petiolare Kunth X X H
Apiaceae
Bowlesia palmata Ruiz & Pav. X H
Cyclospermum laciniatum (DC.) Constance X H
Cyclospermum leptophyllum (Pers.) Sprague X H
Apocynaceae
Sarcostemma solanoides (Kunth) Decne. X Tr
Asteraceae
Achyrocline alata (Kunth) DC. X H
Acmella oleracea (L.) R.K. Jansen X X H
Baccharis salicifolia (Ruiz & Pav.) Pers. X Ab
Chionopappus benthamii S.F. Blake X Ab
Cotula australis (Sieber ex Spreng.) Hook. f. X X H
Erigeron leptorhizon DC. X X H
Galinsoga quadriradiata Ruiz & Pav.r X H
Gamochaeta americana (Mill.) Wedd. X X H
Lactuca scariola L. X H
Ophryosporus peruvianus (J.F. Gmel.) R.M. King & H. Rob. X X Ab
Ophryosporus pubescens (Sm.) R.M. King & H. Rob. X Ab
Philoglossa peruviana DC. X X H
Talavera C., P. Jimnez, A. Ortega, L. Villegas y F. Villasante.
2001. Productividad primaria de las plantas anuales en
las lomas de Meja (Mollendo-Arequipa), en relacin con
la infuencia del fenmeno El Nio Arequipa-Per, en
Seminario Taller: Impacto Biolgico de los eventos El
Nio, Consejo Nacional de Ciencia y Tecnologa (CON-
CYTEC) Lima-Per. pp: 267-270.
Tovar O. 1993. Las Gramneas (Poceas) del Per. Ruizia. Tomo
13. Madrid. Pp. 481.
Apndice 1. Especies registradas por taxa superior, localidad y forma de crecimiento en las lomas de Villa Mara y Amancaes, setiembre-
octubre 2011. (Ar): rbol; (Ab): Arbusto; (H): Hierba; (Su): Suculenta; (Tr): Hierba Trepadora.
(Contina...)
155
Flora vascular de las lomas de Villa Mara y Amancaes
Rev. peru. biol. 19(2): 149 - 158 (August 2012)
Senecio abadianus DC. X Ab
Siegesbeckia fosculosa L'Hr. X X H
Sonchus oleraceus L. X X H
Trixis cacalioides (Kunth) D. Don X Ab
Villanova oppositifolia Lag. X X H
Begoniaceae
Begonia octopetala L'Hr. X X H
Boraginaceae
Cryptantha limensis I.M. Johnst. X H
Cryptantha parvifora Reiche X H
Heliotropium angiospermum Murray X H
Heliotropium arborescens L. X X Ab
Heliotropium indicum L. X H
Heliotropium pilosum Ruiz & Pav. X H
Heliotropium rufpilum (Benth.) I.M. Johnst. X H
Nama dichotoma (Ruiz & Pav.) Choisy X H
Brassicaceae
Capsella bursa-pastoris (L.) Medik. X H
Lepidium virginicum L. X H
Sisymbrium orientale L. X H
Cactaceae
Cleistocactus acanthurus (Vaupel) D.R. Hunt X X Su
Cleistocactus sextonianus (Backeb.) D.R. Hunt X Su
Haageocereus limensis (Salm-Dyck) F. Ritter X X Su
Haageocereus multangularis (Haw.) F. Ritter X Su
Calceolariaceae
Calceolaria angustifora Ruiz & Pav. X Ab
Calceolaria pinnata L. X X H
Caprifoliaceae
Astrephia chaerophylloides (Sm.) DC. X X H
Valeriana pinnatifda Ruiz & Pav. X H
Caricaceae
Vasconcellea candicans (A. Gray) A. DC. X Ar
Caryophyllaceae
Cerastium glomeratum Thuill. X X H
Drymaria paposana var. weberbaueri (Muschl.) Duke X H
Polycarpon tetraphyllum (L.) L. X H
Sagina apetala Ard. X H
Silene gallica L. X H
Stellaria media (L.) Vill. X X H
Convolvulaceae
Ipomoea dumetorum Willd. ex Roem. & Schult. X Tr
Jacquemontia unilateralis (Roem. & Schult.) O'Donell X Tr
Crassulaceae
Crassula connata (Ruiz & Pav.) A. Berger & al. X X H
Cucurbitaceae
Cyclanthera mathewsii Arn. X Tr
Cyclanthera sp X Tr
Sicyos baderoa Hook. & Arn. X X Tr
Euphorbiaceae
Andrachne microphylla (Lam.) Baill. X H
Chamaesyce hypericifolia (L.) Millsp. X H
Chamaesyce serpens (Kunth) Small X H
Cnidoscolus basiacanthus (Pax & K. Hoffm.) J.F. Macbr. X Ab
Croton alnifolius Lam. X Ab
Ricinus communis L. X Ab
Fabaceae
Vachellia macracantha (Humb. & Bonpl. ex Willd.) Seigler & Ebinger X Ar
Caesalpinia spinosa (Molina) Kuntze X X Ar
Taxa superior Especie Villa Mara Amancaes
Forma de
crecimiento
Apndice 1. Continuacin.
(Contina...)
156
Trinidad et al.
Rev. peru. biol. 19(2): 149 - 158 (Agosto 2012)
Crotalaria incana L. X H
Hoffmannseggia prostrata Lagerh. ex DC. X H
Medicago polymorpha L. X H
Parkinsonia aculeata L. X Ar
Geraniaceae
Erodium cicutarium (L.) L'Hr. ex Aiton X H
Erodium malacoides (L.) L'Hr. ex Aiton X X H
Erodium moschatum (L.) L'Hr. ex Aiton X X H
Lamiaceae
Hyptis sidifolia (L'Hr.) Briq. X H
Marrubium vulgare L. X H
Salvia paposana Phil. X X H
Salvia rhombifolia Ruiz & Pav. X X H
Stachys arvensis L. X H
Loasaceae
Loasa nitida Desr. X X H
Mentzelia scabra subsp. grandifora (Ruiz & Pav. ex G. Don) Weigend X Ab
Nasa urens (Jacq.) Weigend X X H
Malvaceae
Fuertesimalva chilensis (A. Braun & C.D. Bouch) Fryxell X X H
Fuertesimalva peruviana (L.) Fryxell X X H
Montiaceae
Calandrinia alba (Ruiz & Pav.) DC. X X H
Cistanthe lingulata (Ruiz & Pav.) Hershkovitz X H
Cistanthe paniculata (Ruiz & Pav.) Carolin ex Hershkovitz X H
Nyctaginaceae
Mirabilis prostrata (Ruiz & Pav.) Heimerl X X H
Oxalidaceae
Oxalis lomana Diels X H
Oxalis megalorrhiza Jacq. X X H
Papaveraceae
Fumaria capreolata L. X X H
Plantaginaceae
Plantago major L. X H
Veronica persica Poir. X H
Plumbaginaceae
Plumbago coerulea Kunth X H
Polygonaceae
Emex spinosa (L.) Campd. X H
Primulaceae
Anagallis arvensis L. X H
Solanaceae
Cestrum auriculatum L'Hr. X Ab
Exodeconus prostratus (L'Hr.) Raf. X H
Nicotiana paniculata L. X X H
Nolana humifusa (Gouan) I.M. Johnst. X X H
Solanum montanum L. X X H
Solanum peruvianum L. X X H
Urticaceae
Parietaria debilis G. Forst. X X H
Urtica urens L. X X H
Verbenaceae
Lantana scabiosifora Kunth X H
Verbena litoralis Kunth X H
Polypodiopsida
Pteridaceae
Adiantum poiretii Wikstr. X H
Taxa superior Especie Villa Mara Amancaes
Forma de
crecimiento
Apndice 1. Continuacin.
157
Flora vascular de las lomas de Villa Mara y Amancaes
Rev. peru. biol. 19(2): 149 - 158 (August 2012)
Apndice 2. Especies frecuentes en las lomas de Villa Mara y Amancaes. (a)Cleistocactus acanthurus (Cactaceae); (b)Anthericum ec-
cremorrhizum (Asparagaceae); (c)Nasa urens (Loasaceae); (d)Alstroemeria lineatifora (Alstroemeriaceae); (e)Sarcostemma solanoides
(Apocynaceae); (f)Drymaria paposana var. weberbaueri (Caryophyllaceae); (g)Exodeconus prostratus (Solanaceae);(h)Solanum montanum
(Solanaceae); (i)Jacquemontia unilateralis (Convolvulaceae); (j)Loasa nitida (Loasaceae); (k)Begonia octopetala (Begoniaceae); (l)Erigeron
leptorhizon (Asteraceae); (m)Senecio abadianus (Asteraceae); (n)Salvia paposana (Lamiaceae); (o)Chionopappus benthamii (Asteraceae).
158
Trinidad et al.
Rev. peru. biol. 19(2): 149 - 158 (Agosto 2012)
Apndice 3. Especies frecuentes en las lomas de Villa Mara y Amancaes. (a) Oziroe bifora (Asparagaceae); (b) Cyclanthera mathewsii
(Cucurbitaceae); (c)Heliotropium rufpilum (Boraginaceae); (d)Oxalis lomana (Oxalidaceae); (e) Lamarckia aurea (Poaceae); (f)Philoglosa
peruviana (Asteraceae); (g)Fuertesimalva peruviana (Malvaceae); (h) Erodium malacoides (Geraniaceae); (i)Nolana humifusa (Solanaceae);
(j)Calceolaria pinnata (Calceolariaceae); (k) Villanova oppositifolia (Asteraceae); (l)Acmella oleracea (Asteraceae); (m)Commelina fasciculata
(Commelinaceae); (n) Croton alnifolius (Euphorbiaceae); (o)Galinsoga quadriradiata (Asteraceae) .
159
Dieta de murcilagos filostmidos de Cusco - Per
Rev. peru. biol. 19(2): 159 - 166 (August 2012)
Rev. peru. biol. 19(2): 159 - 166 (Agosto 2012)
Facultad de Ciencias Biolgicas UNMSM
ISSN 1561-0837
Dieta de murcilagos flostmidos del valle de Kosipata, San Pedro,
Cusco - Per
Rossana Maguia
1
, Jessica Amanzo
2
y Luis Huamn
1
Diet of phyllostomid bats of Kosipata Valley, San Pedro, Cusco - Peru
1 Laboratorio de Palinologa y
Paleobotnica. Laboratorios de
Investigacin y Desarrollo (LID).
Universidad Peruana Cayetano
Heredia. Av. Honorio Delgado
430, Urb. Ingeniera, S.M.P. Lima
31, Per.
2 Laboratorio de Estudios en Bio-
diversidad. Laboratorios de Inves-
tigacin y Desarrollo (LID). Univer-
sidad Peruana Cayetano Heredia.
Email Rossana Maguia:
nrmaguina@gmail.com
Presentado: 13/12/2011
Aceptado: 12/08/2012
Publicado online: 10/10/2012
Resumen
En el presente estudio se evalu la dieta de murcilagos flostmidos de los bosques montanos del valle de
Kosipata (San Pedro, Cusco). Las especies evaluadas pertenecieron a los gneros Carollia, Sturnira y Anoura.
Se analiz la dieta en base a la presencia de polen y semillas en muestras fecales. Se realiz un anlisis de la
importancia de las plantas consumidas, la amplitud del nicho alimenticio y el nivel de superposicin del nicho
alimenticio. Los resultados mostraron que la dieta de los murcilagos est compuesta en su mayora por frutos
y nctar-polen de las plantas. Los principales recursos registrados pertenecieron a los gneros Piper (registrado
en 23,08% de las muestras), Cecropia (en 21,15%) y Abutilon (en 13,46%), los cuales fueron los recursos ms
importantes para la comunidad de murcilagos evaluada. La amplitud de nicho alimenticio para las 6 especies
analizadas indic que son especialistas en su dieta en el rea de estudio. Por otro lado, los mayores niveles
de superposicin de nicho alimenticio ocurre entre las especies C. brevicauda, C. perspicillata y S. erythromos.
Palabras clave: Phyllostomidae; murcilago; frugvoro; nectarvoro; dieta.
Abstract
In this study had been evaluated the diet of phyllostomids bats of the montane forest of the Kosipata Valley
(San Pedro, Cusco). The species of bats evaluated belong to the genus Carollia, Sturnira and Anoura. The
diet was evaluated regarding the presence of pollen and seeds in fecal samples. Also, it was analyzed the
importance of the plants consumed, the food niche breath and the food niche overlap. The results showed
that the diet of the bats is mostly compounded of fruit and nectar-pollen of the plants. The principal resources
registered belong to the genus Piper (registered in 23,08% of the samples), Cecropia (in 21,15%) and Abutilon
(in 13,46%), which was the most important resources for the community of bats evaluated. The food niche
breath of the six species analyzed showed that they are specialist in their diet in the study zone. The highest
level of food niche overlap occurs between the species C. brevicauda, C. perspicillata and S. erythromos.
Keywords: Phyllostomidae; bat; frugivorous; nectarivorous; diet.
Introduccin
En el Per, las investigaciones realizadas en el orden Chi-
roptera han servido para llenar vacos de informacin sobre
distribucin, diversidad y sistemtica de especies. Aunque la
mayora de ellos incluye aspectos ecolgicos, son muy pocos los
dedicados al estudio exclusivo de la dieta (Novoa et al. 2011,
Arias et al. 2009, Gorchov et al. 1995, Ascorra et al. 1996).
La importancia de los estudios de dieta en murcilagos, espe-
cfcamente en especies herbvoras, radica en los roles ecolgicos
de polinizacin y dispersin que cumplen dentro de su hbitat
(Fleming 1988). Los murcilagos polinizadores son importan-
tes en la fenologa reproductiva y la estructura poblacional de
las plantas, mientras que los dispersores se convierten en entes
esenciales de la regeneracin de bosques tropicales y del man-
tenimiento de la diversidad vegetal del mismo (Fleming 1988).
En el presente estudio, (1) se identifcan los taxa componentes
de la dieta del murcilagos frugvoros Carollia y Sturnira (Gard-
ner 2007), y del gnero nectarvoro Anoura (Ortega y Alarcn
2007); y (2) se determinan los taxa ms importantes en la dieta
y realiza un anlisis del nicho alimenticio, considerando la am-
plitud y superposicin de nicho, con la fnalidad de documentar
las relaciones existentes entre los murcilagos y las plantas en
el bosque nublado del valle de Kosipata, San Pedro, Cusco.
Material y mtodos
rea de estudio.- El estudio se realiz en el Valle de Kosipa-
ta, en la localidad de San Pedro (Cusco), en un rea circundante
al alberge turstico Cock of the Rocks. La zona se encuentra
localizada en los bosques montano orientales del sureste amaz-
nico, abarcando altitudes desde 1100 hasta 1600 m. Comprende
las siguientes zonas de vida segn el sistema de clasifcacin de
Holdrige (1978): Bosque muy Hmedo Montano Subtropical
(bmh-MS), Bosque muy Hmedo Montano Bajo Subtropical
(bmh-MBS) y Bosque Pluvial Montano Subtropical (bp-MS).
El clima es templado, se ha reportado que dependiendo de la
altura, el rango de temperatura vara entre 8 y 18 C, con una
precipitacin anual de 4510 a 5470 mm (SENAMHI 2010). La
humedad relativa es alta durante todo el ao, pero se observan
dos pocas climticas marcadas; la poca de lluvias, que va de
diciembre a abril, y la poca seca, de mayo a noviembre, con
escazas lluvias. Pacheco et al., (2004) describen la localidad de
San Pedro como un rea dominada por bosque secundario, con
laderas moderadas de 20 30, y varios riachuelos pequeos
que alimentan un largo torrente con fujo hacia el este. Adems
mencionan que el paisaje est dominado por gruesas plantas
de bamb trepador (Chusquea sp.), y por rboles cubiertos por
epftas. El acceso hacia esta zona se realiza desde la ciudad de
Cusco, va terrestre, por la carretera a Pilcopata.
Toma de datos.- Se llev a cabo durante la poca seca en
julio del 2007 y la poca de lluvias en marzo y abril del 2008.
Las muestras fecales de los murcilagos proceden de los indi-
viduos capturados durante el estudio de Delgado (2010). Las
muestras fueron conservadas en alcohol al 70%. Las especies de
murcilagos fueron determinadas usando claves taxonmicas
especializadas (Gardner 2007, Angulo et al. 2004, Albuja 1999,
160
Maguia
Rev. peru. biol. 19(2): 159 - 166 (Agosto 2012)
Especie Gremio alimenticio N
Frecuencia de los componentes
E
P S TV I
Carollia brevicauda frugvoro 10 6 1 9 1 0
Carollia perspicillata frugvoro 3 2 2 3 0 0
Carollia manu frugvoro 1 0 0 1 0 0
Sturnira erythromos frugvoro 9 8 3 5 0 1
Sturnira oporaphilum frugvoro 19 8 2 16 1 2
Sturnira tildae frugvoro 2 1 1 2 0 0
Sturnira lilium frugvoro 1 0 0 1 0 0
Anoura caudifer nectarvoro 6 5 0 4 0 0
Anoura geoffroyi nectarvoro 1 0 0 1 0 0
Total 52 30 (57,69%) 9 (17,3%) 42 (80,76%) 2 (3,84%) 3 (5,76%)
Tabla 1. Frecuencia de los componentes registrados en las muestras fecales. Nmero de muestras (N), polen (P), semilla (S), tejido vegetal
(TV), restos de insectos (I), muestra estril (E).
Pacheco & Solari 1997) o fueron confrontados con la coleccin
de Mamferos del Departamento de Mastozoologa del Museo
de Historia Natural de la Universidad Nacional Mayor de San
Marcos (UNMSM). Tambin se realiz una colecta selectiva de
plantas en foracin o fructifcacin y se elaboraron catlogos
referenciales de semillas y granos de polen. Las muestras bo-
tnicas fueron determinadas usando claves taxonmicas (Pen-
nington et al. 2004, Brako & Zarucchi 1993, Gentry 1993) y
el apoyo de especialistas. Las determinaciones se corroboraron
en el Herbario del Museo de Historia Natural de la UNMSM.
El material fue depositado en la palinoteca del Laboratorio de
Palinologa y Paleobotnica y en el Herbario Magdalena Pavlich
de la Universidad Peruana Cayetano Heredia.
Anlisis de las muestras fecales.- Los anlisis se realiza-
ron sin conocer a que especie de murcilago perteneca cada
muestra. Primero se registraron semillas, tejidos vegetales y
restos de insectos, mediante observaciones con estereoscopio.
Las semillas registradas fueron determinadas utilizando la
gua de Martin y Barkley (1961) y el catlogo referencial.
Seguidamente, las muestras fueron sometidas al proceso de de
acetlisis de Erdtman (1960) modifcado y el material obte-
nido fue montado por duplicado con lminas portaobjetos y
cubreobjetos, y fueron observadas al microscopio (40X). Los
granos de polen registrados se determinaron con las claves
taxonmicas de Roubik y Moreno (1991), Heusser (1971),
Herrera y Urrego (1996) y el catlogo referencial. Finalmente
los resultados se clasifcaron en cuatro categoras: a) semillas,
b) granos de polen, c) tejido vegetal y d) insectos. Se elabor
una lista de plantas consumidas en base a los resultados de
semillas y granos de polen.
ndice Valor de Importancia de las Plantas (IVIP).- Se cal-
cul utilizando las frecuencias de ocurrencia de granos de polen
y semillas (planteado por Amaya 1991 citado en Muoz-Saba
et al. 1997). Entre mayor es el IVIP, mayor es el uso que una
especie o comunidad de murcilagos da a una planta.
Anlisis del nicho alimenticio.- En base a los resultados
de granos de polen se estim la amplitud de nicho alimenticio
utilizando la medida de Levins estandarizada y la superposi-
cin de nicho alimenticio utilizando el ndice de MacArthur
y Levins modifcado por Pianka (Krebs 1999). Estos ndices
fueron validados con la prueba estadstica de Kruskal-Wallis
(Lpez & Vaughan 2007). Adems, sometimos los resultados
de superposicin de nicho a un anlisis de agrupamiento. Para
los anlisis estadsticos se utiliz el programa PAST (Hammer
et al. 2001).
Resultados
Anlisis de las muestras fecales.- Se analizaron 52 muestras
fecales pertenecientes a 9 especies de murcilagos de la familia
Phyllostomidae: Carollia brevicauda, C. perspicillata, C. manu,
Sturnira erythromos, S. oporaphilum, S. tildae, S. lilium, Anoura
caudifer y A. geofroyi. La mayora de las muestras contenan
tejidos vegetales (80,7%, n= 42) y granos de polen (57,6%, n=
30). Tambin se registraron 3 muestras estriles (Snchez-Casas
& lvarez 2000), es decir aquellas que no contenan ninguno
de los 4 componentes analizados (Tabla 1).
En las muestras fecales se registraron 79 morfotipos de granos
de polen y 9 morfoespecies de semillas. Los principales recursos
consumidos pertenecieron a los gneros Piper (registrado en
23,08% de las muestras), Cecropia (en 21,15%) y Abutilon (en
13,46%) (Apndice 1). Las especies C. brevicauda, C. perspici-
llata, S. erythromos y S. oporaphilum consumieron en su mayora
especies de los gneros Piper y Cecropia, adems S. erythromos
consumi varias especies de la familia Solanaceae; mientras que
A. caudifer consumi mayormente especies del gnero Abutilon.
En las muestras fecales de C. manu, S. lilium y A. geofroyi no se
registraron los mencionados componentes.
ndice de Valor de Importancia de las plantas (IVIP).-
Los resultados del IVIP mostraron que para la comunidad de
murcilagos evaluados en el valle de Kosipata las plantas ms
importantes pertenecen a los gneros Piper, Cecropia y Abutilon,
con 0,675, 0,650 y 0,385 de IVIP respectivamente (Tabla 2).
Las plantas del gnero Piper son las ms importantes para las
especies C. brevicauda, C. perspicillata y S. erythromos, las plantas
de Cecropia son las ms importantes para S. oporaphilum, y las
plantas de Abutilon para la especie A. caudifer.
Anlisis del nicho alimenticio.- Los resultados de amplitud
de nicho alimenticio segn la medida de Levins estandarizada (B
A
)
mostraron valores cercanos a cero para las 6 especies analizadas,
indicando que son especialistas en su dieta en el rea de estudio
161
Dieta de murcilagos filostmidos de Cusco - Per
Rev. peru. biol. 19(2): 159 - 166 (August 2012)
Familia Gnero
Cb Cp Se So St Ac
IVIP*
P P P P P P
Piperaceae Piper spp. 0,154 0,200 0,250 0,071 - - 0,675
Cecropiaceae Cecropia spp. 0,077 0,100 0,125 0,107 0,167 0,074 0,650
Malvaceae Abutilon spp. - 0,200 - - - 0,185 0,385
Flacourtiaceae Flacourtiaceae sp1. 0,038 - 0,125 - - 0,111 0,275
Melastomataceae Miconia sp. - - - 0,036 0,167 0,037 0,239
Solanaceae cf. Solanum sp. - - - - 0,167 0,037 0,204
Euphorbiaceae cf Omphalea sp. - - - 0,036 0,167 - 0,202
Asteraceae Mikania sp. - - 0,125 0,071 - - 0,196
Rosaceae Rubus spp. 0,038 0,100 - - - 0,037 0,175
Moraceae-Urticaceae Moraceae-Urticaceae sp4. - - - - 0,167 - 0,167
Dicotiledonea - - - - 0,167 - 0,167
Cucurbitaceae Cayaponia sp. - - - 0,036 - 0,111 0,147
Rubiaceae Psychotria spp. 0,038 - - 0,071 - 0,037 0,147
Heliconiaceae Heliconia sp. 0,038 0,100 - - - - 0,138
Bromeliaceae Pitcairnia paniculata - 0,100 - - - 0,037 0,137
Bromeliaceae Vriesea sp. - 0,100 - - - 0,037 0,137
Solanaceae Solanum-Lycopersicum 0,038 - 0,063 0,036 - - 0,137
Solanaceae Solanaceae sp2. - - 0,125 - - - 0,125
Gentianaceae cf. Chelonanthus sp. - - - 0,036 - 0,074 0,110
Ulmaceae cf Celtis sp. 0,038 - 0,063 - - - 0,101
Moraceae-Urticaceae Moraceae-Urticaceae sp2. - 0,100 - - - - 0,100
cf Ulmaceae - 0,100 - - - - 0,100
Indeterminada 6 - 0,100 - - - - 0,100
Tabla 2. ndice de Valor de Importancia de las plantas consumidas por los murcilagos. C. brevicauda (Cb), C. perspicillata (Cp), S. erythromos
(Se), S. oporaphilum (So), S. tildae (St), A. caudifer (Ac), uso (P), ndice de valor de importancia de las plantas (IVIP). * Slo se muestran los
resultados de IVIP mayores a 0,1.
Especie N B BA
Carollia brevicauda 6 1,1143 0,0014
Carollia perspicillata 2 1,0577 0,0007
Sturnira erythromos 8 1,0335 0,0004
Sturnira oporaphilum 6 1,9371 0,0119
Sturnira tildae 1 3,9032 0,0367
Anoura caudifer 5 4,5518 0,0450
Tabla 3. ndice de Levins estandarizado (B
A
). Nmero de muestras
(N), medida de Levins (B).
Figura 1. Anlisis de agrupamiento. Abreviaciones: C. brevicauda
(Cb), C. perspicillata (Cp), S. erythromos (Se), S. oporaphilum (So),
S. tildae (St), A. caudifer (Ac).
0 0,2 0,4 0,6 0,8 1,0
Indice de Pianka
St
Ac
So
Se
Cb
Cp
(Tabla 3). La prueba de Kruskal-Wallis indic que no existen di-
ferencias signifcativas entre ellos (p> 0,05; H= 6,232, p= 0,397).
Los resultados del ndice de MacArthur y Levins modifcado
por Pianka mostraron que existe una superposicin casi completa
entre la dieta de las especies C. brevicauda, C. perspicillata y S.
erythromos (con 0,9999); mientras que los restantes niveles de
superposicin son todos menores a 0,5. La prueba de Kruskal-
Wallis indic que existen diferencias signifcativas entre dichos
resultados (p < 0,05, H = 30,95, p = 0,005). El anlisis de
agrupamiento result en un dendrograma que agrupa a 3 de las
especies analizadas, C. brevicauda, C. perspicillata y S. erythromos.
S. oporaphilum muestra una dieta ligeramente similar al grupo
anteriormente mencionado, mientras que S. tildae y A. caudifer
presentan dietas dismiles a las dems especies (Fig. 1).
Discusin
Dieta.- La dieta de los murcilagos evaluados estuvo repre-
sentada en su mayora por las familias y gneros siguientes: Pi-
peraceae (Piper), Cecropiaceae (Cecropia), Malvaceae (Abutilon)
y Solanaceae. Todas ellas registradas como parte de la dieta de
otras comunidades de murcilagos en el Neotrpico (Novoa et
al. 2011, Arias et al. 2009, Lou & Yurrita 2005, Ties & Kalko
2004, Lpez & Vaughan 2004, Muchhala & Jarrn-V 2002,
Giannini 1999, Ascorra et al. 1996, Gorchov et al. 1995). Las
162
Maguia
Rev. peru. biol. 19(2): 159 - 166 (Agosto 2012)
familias utilizadas para el consumo de frutos presentan frutos
de tipo nuez o drupa ssiles en infrutescencias de tipo espiga
(Cecropia y Piper) (Ties & Kalko 2004, Gentry 1993), o de tipo
baya como en el caso de Solanaceae (Gentry 1993). La familia
Malvaceae, gnero Abutilon, que fue utilizada para el consumo
de nctar-polen presenta algunas caractersticas que coinciden
con las descritas para las plantas con sndrome de quiroptero-
flia (Muchhala & Jarrn-V 2002), tales como, fores solitarias
o agrupadas, con corola acampanada de colores claros, muchas
veces en ramas terminales de fcil acceso para los murcilagos
(Pennington et al. 2004).
Cada especie de murcilago mostr un mayor consumo hacia
ciertos gneros o especies de plantas. Las especies del gnero
Carollia mostraron un mayor consumo hacia plantas del gnero
Piper. En las especies de Sturnira se observaron preferencias
variadas, S. erythromos consumi mayormente plantas de Piper
y varios taxa de la familia Solanaceae; mientras que S. opora-
philum consumi preferentemente plantas de Cecropia y Piper.
En el caso de A. caudifer se observ un mayor consumo hacia
plantas del gnero Abutilon. Estos resultados concuerdan con
lo observado por otros autores para las especies de murcilagos
evaluadas (Ortega & Alarcon 2007, Lou & Yurrita 2005, Ties
& Kalko 2004, Lpez & Vaughan 2004, Muchhala & Jarrn-V
2002, Giannini 1999).
Adems, los resultados obtenidos apoyan la idea de que los
murcilagos frecuentemente tienden a consumir frutos de una
o dos especies de plantas y complementan su dieta con otros
recursos menos importantes para ellos (Lou & Yurrita 2005,
Lpez 1996). As como tambin, el registro del consumo de
frutos de plantas pertenecientes a gneros o familias que con-
tienen especies pioneras (Piper, Cecropia y la familia Solanaceae)
(Ties & Kalko 2004, Lobova et al. 2003, Olea-Wagner et al.
2007), apoya la importancia de los murcilagos frugvoros en
los procesos de manutencin de los bosques (Novoa et al. 2011,
Fleming & Heithaus 1981).
Las especies de los gneros Carollia y Sturnira analizadas, no
son estrictamente frugvoras, ya que se ha registrado la ingesta
de nctar-polen e insectos, aunque en menor cantidad que la
ingesta de frutos (registrado por medio de tejidos vegetales y
semillas). Del mismo modo, la especie A. caudifer tampoco es
estrictamente nectarvora, debido al registro de tejidos vegetales
en las muestras y a la determinacin del uso de frutos de algunos
taxa (Apndice 1).
ndice de Valor de Importancia de las Plantas (IVIP): La
importancia de los gneros Piper y Cecropia en la dieta de mur-
cilagos frugvoros ha sido previamente recalcada por diversos
autores (Lou & Yurrita 2005, Gonalves Da Silva et al. 2008,
Mello et al. 2004, Lpez & Vaughan 2007, Ties & Kalko 2004,
Lobova et al. 2003, Giannini 1999), guardando concordancia
con los resultados del IVIP (Tabla 2). Sin embargo, se contra-
ponen a aquellos estudios que consideran a Cecropia como un
complemento de la dieta de estos mamferos (Mello et al. 2008,
2004, Muoz-Saba et al. 1997).
La gran importancia del gnero Piper puede explicarse por
el tipo de fenologa que posee. Las inforescencias contienen
algunos frutos maduros en intervalos de tiempo predecibles
durante largos periodos, fenologa conocida como estado-estable;
adems presentan una maduracin de frutos nocturna (Ties &
Kalko 2004). En el caso del gnero Cecropia, puede deberse a su
gran abundancia en la zona de estudio (obs. pers), ofreciendo
considerables cantidades de frutos. Lo cual coincide con Dumont
(2003), quien mencion que Cecropia es relativamente abun-
dante en las zonas tropicales y produce frutos constantemente.
El gnero Abutilon tambin ha sido mencionado como una
fuente importante de nctar-polen para murcilagos nectarvo-
ros neotropicales en los trabajos de Arias et al. (2009) y Sazima
et al. (1999), lo cual concuerda con nuestros resultados. La
importancia de estas plantas podra ser resultado del patrn de
foracin que presentan, como menciona Sazima et al. (1999)
para la especie Abutilon af. regnellii, presenta un patrn de
foracin continuo, ofreciendo nctar-polen a lo largo del ao.
Sin embargo, es necesario realizar estudios fenolgicos de las
especies de Abutilon de Kosipata para tener certeza del patrn
de foracin que presentan.
Otra familia que es ampliamente mencionada en la literatura
como utilizada por murcilagos del gnero Sturnira es Solanaceae
(Giannini 1999, Mello et al. 2008, Lou & Yurrita 2005, Lpez
& Vaughan 2007), lo cual concuerda con nuestros resultados
para la especie S. erythromos. Para los taxa de dicha familia se
obtuvieron bajos IVIP, pero al calcular la sumatoria de los IVIP
de los 4 taxa, el IVIP general para la familia Solanaceae pasa a
ocupar el tercer lugar en la tabla de resultados con 0,528; lo
cual realza la importancia de dicha familia en la dieta de los
murcilagos del gnero Sturnira.
Anlisis del nicho alimenticio.- El ndice de Levins estan-
darizado (B
A
) calculado indica que las especies C. brevicauda, C.
perspicillata, S. erythromos, S. oporaphilum, S. tildae y A. caudifer
son especialistas en su dieta en la zona de estudio. Los resultados
de Lou y Yurrita (2005) sobre la amplitud de nicho alimenticio
muestran que las especies C. brevicauda, C. perspicillata y S.
lilium son bastante selectivas (especialistas). Igualmente, Lpez y
Vaughan (2007) obtuvieron bajos B
A
para 3 especies de Carollia.
Sobre las especies del gnero Anoura, se menciona en trabajos
previos, que pueden consumir una mayor variedad de recursos
que las especies frugvoras (Caballero-Martnez et al. 2009,
Arias et al. 2009), sin embargo no se han encontrado datos de
amplitud de nicho alimenticio que indiquen si son especialistas
o generalistas en sus dietas. Dentro del gremio de nectarvoros
existen especies especialistas como Glossophaga soricina (Lou &
Yurrita 2005), por lo tanto no se descarta la posibilidad de que A.
caudifer pueda tener un comportamiento selectivo al momento
de consumir sus alimentos, pero es necesario estudios posteriores
que analicen un mayor nmero de muestras fecales.
La prueba de Kruskal-Wallis para los resultados de B
A
indic
que no existen diferencias signifcativas entre ellos (p> 0,05;
H= 6,232, p= 0,397), es decir, no hay una especie que sea ms
especialista en su dieta que otra en Kosipata. Sin embargo, dada
la variedad de registros en las muestras fecales de la especie A.
caudifer, se considera que dicha especie sera menos especialista
que las dems especies analizadas, pero para comprobar dicha
afrmacin es necesario el anlisis de un mayor nmero de
muestras.
Los resultados del ndice de MacArthur y Levins modifcado
por Pianka mostraron que existe superposicin entre las dietas
de las especies C. brevicauda, C. perspicillata y S. erythromos
(Tabla 4). Estos resultados coinciden con Lpez (1996) (citado
163
Dieta de murcilagos filostmidos de Cusco - Per
Rev. peru. biol. 19(2): 159 - 166 (August 2012)
en Lou & Yurrita 2005), Lpez y Vaughan (2007) y Lou y
Yurrita (2005), quienes mencionan que existen altos niveles de
superposicin entre las dietas de especies de Carollia, Sturnira
y otros Stenodermatinos. Morfolgicamente las especies de
Carollia y Sturnira son de tallas similares, lo cual origina mayor
superposicin de nicho trfco, ya que los frutos que consumen
por lo general son de tamaos similares (Lou & Yurrita 2005).
En conclusin, se ha logrado documentar las interacciones
que existen entre los murcilagos y las plantas en el valle de
Kosipata, evidenciadas por el uso que los murcilagos hacen de
los frutos y el nctar-polen como parte de su dieta. Cabe resaltar
que estos murcilagos podran estar desempeando importantes
roles de dispersin y polinizacin, siendo entes fundamentales
en los procesos de sucesin y manutencin del bosque.
Agradecimientos
Nuestro especial agradecimiento al Blgo. Daniel Blanco y a los
trabajadores del albergue turstico Cock of the rocks. Al PhD.
Vctor Pacheco, Director del Departamento de Mastozoologa
del Museo de Historia Natural (UNMSM), por la revisin y
correcciones al presente trabajo. A Carla Delgado, Alessandra
Quiones, Cindy Vergel, Aaron Cnepa, Jorge Len, Daniel
Ramos y Alfredo Butrn, por su participacin en la colecta de
muestras en Kosipata.
Literatura citada
Albuja L. 1999. Quirpteros del Ecuador. Editorial Cicetronic Cia.
Ltda. Offset. Quito-Ecuador; 2da edicin.
Angulo S.R, Escobedo M, Lpez C & Ros J.A. 2004. Introduccin
al estudio y conservacin de los Quirpteros Neotropi-
cales. Universidad Nacional de la Amazona Peruana.
Iquitos, Per.
Arias E., Cadenillas R., Pacheco V. 2009. Dieta de murcilagos
nectarvoros del Parque Nacional Cerros de Amotape,
Tumbes. Rev. peru. biol. 16(2): 187 190.
Ascorra C.F., Solari S. & Wilson D. E. 1996. Diversidad y ecologa
de los quirpteros en Pakitza. Pp. 585-604. En Manu:
the Biodiversity of southeastern Peru (d.E. Wilson & A.
Sandoval, eds.). Smithsonian Institution, Washington, DC.
Brako L. & Zarucchi J. L. 1993. Catlogo de las angiospermas y
gimnospermas del Per. Monographs in Sistematic Botany.
Missouri Botanical Garden. Vol. 45.
Caballero-Martinez L. A., Rivas I. V. & Aguilera L. I. 2009. Hbitos
alimentarios de Anoura geoffroyi (Chiroptera: Phyllos-
tomidae) en Ixtapan del Oro, Estado de Mxico. Acta
Zoolgica Mexica (n. s.) 25(1): 161 175.
Delgado C. 2010. Variacin estacional en la diversidad y estado
reproductivo de quirpteros con nfasis en los gneros
Sturnira y Carollia (Chiroptera: Phyllostomidae) en el
bosque nublado de San Pedro, Cusco (Tesis). Universidad
Peruana Cayetano Heredia.
Dumont E. R. 2003. Bats and fruit: an ecomorphological approach.
En T. H. Kunz and M. B. Fenton (Eds). Bat Ecology,
Pp. 398-421. The University of Chicago Press, Chicago,
Illinois.
Erdtman G. 1960. The acetolysis method. Svensk. Bot Tidshkr.
54(4): 561 564.
Fleming T. H. 1988. The short-tailed fruit bat: a study in plant-
animal interactions. University of Chicago Press, Chicago,
Illinois, USA.
Fleming T. H. & Heithaus E. R. 1981. Fugivorous bats, seed shadows,
and the structure of Tropical Forests. Biotropica, Vol. 13,
No. 2, Supplement: Reproductive Botany 45 43.
Gardner A. L. (Ed.) 2007. Mammals of South America. Volume 1:
Marsupials, Xenarthrans, Shrews and Bats. USA.
Gentry A. 1993. A feld guide to the families and genera of Woody
Plants of Northwest South America (Colombia, Ecuador,
Peru) with supplementary notes on herbaceous taxa. Con-
servation International. Washington, DC. USA.
Giannini N. P. 1999. Selection of diet and elevation by sympatric
species os Sturnira in an Andean Rainforest. Journal of
Mammology 80(4): 1186 1195.
Gonalves Da Silva A., Gaona A. & Medellin R. 2008. Diet and Tro-
phic structure in a community of fruit-eating bats in Lacan-
don Forest, Mexico. Journal of Mammology 89(1): 43 49.
Gorchov D. L., Cornejo F., Ascorra C. F. & JARAMILLO M. 1995.
Dietary overlap between frugivorous birds and bats in the
Peruvian Amazon. Oikos 74: 235 250.
Hammer , Harper, D.A. & RYAN P.D. 2001. PAST: Paleontolo-
gical Statistics Software Package for Education and Data
Analysis. Paleontologia Electronica 4(1).
Herrera L. F. & Urrego L. E. 1996. Atlas de polen de plantas tiles
y cultivadas de la Amazona colombiana. Volumen XL.
Tropenbos. Colombia.
Heusser C. 1971. Pollen and spores of Chile. The University of
Arizona Press. United States of America.
Holdrige L. 1978. Ecologa Basada en Zonas de Vida. IICA. San
Jos - Costa Rica.
Krebs C. J. 1999. Ecological Methodology. Second Edition. Addison-
Wesley Educational Publishers, Inc. Benjamin/Cummings.
Menlo Park, California.
Lobova T. A., Mori S., Blanchard F., Peckham H. & Charles-
Dominique P. 2003. Cecropia as a food resource for bats
in French Guiana and the signifcance of fruit structure in
seed dispersal and longevity. American Journal of Botany
90(3): 388-403.
Loayza P., Rios R. & Larrea-Alcazar D. 2006. Disponibilidad de
recurso y dieta de murcilagos frugvoros en la Estacin
Biolgica de Tunquini, Bolivia. Ecologa en Bolivia
41(1): 723.
Lopez J. E., Vaughan C. 2004. Observations on the role of frugi-
vorous bats as seed dispersers in Costa Rican secondary
humid forests. Acta Chiropterologica 6(1): 111 119.
Lopez J. E. & Vaughan C. 2007. Food Niche Overlap Among Neo-
tropical Frugivorous bats in Costa Rica. Rev. Biol. Trop.
Vol.55(1):301313.
Lou S. & Yurrita C. 2005. Anlisis de nicho alimentario en la
comunidad de murcilagos frugvoros de Yaxh, Petn,
Guatemala. Acta Zoolgica Mexicana (n. s.) 21(1): 83 94.
Martin A. C. & Barkley W. D. 1961. Seed Identifcation Manual. Uni-
versity of California Press. Berkely and Los Angeles, USA.
Mello M. A., Schittini G. M., Selig P. & Bergallo H. G. 2004. Sea-
sonal variation in the diet of the bat Carollia perspicillata
(Chiroptera: Phyllostomidae) in an Atlantic Forest area in
southeastern Brazil. Mammalia 68(1): 4955.
Mello M., Kalko E. & Silva W. 2008. Diet and abundance of the
bat Sturnira lilium (Chiroptera) in a Brazilian Montane
Atlantic Forest. Journal of Mammalogy 89(2): 485492.
Cb Cp Se So St Ac
C. brevicauda - 0,9999 0,9999 0,3528 0,0053 0,0016
C. perspicillata - 0,9999 0,3527 0,0040 0,0065
S. erythromos - 0,3527 0,0009 0,0013
S. oporaphilum - 0,0143 0,0024
S. tildae - 0,0850
A. caudifer -
Tabla 4.- ndice de MacArthur y Levins modifcado por Pianka. C.
brevicauda (Cb), C. perspicillata (Cp), S. erythromos (Se), S. opora-
philum (So), S. tildae (St), A. caudifer (Ac).
164
Maguia
Rev. peru. biol. 19(2): 159 - 166 (Agosto 2012)
Familia Especie Uso
Cb Cp Se So St Ac
F A F A F A F A F A F A
SEMILLAS
Cecropiaceae Cecropia latiloba f 1 3
Clusiaceae Vismia tomentosa f 1 1
Piperaceae
Piper cf crassinervium f 1 2 1 5
Piper sp1. f 1 1
Piper sp2. f 1 25
Piper sp3. f 1 3
Solanaceae Solanum appressum f 1 2
cf. Fabaceae f 1 1
cf. Moraceae f 1 1
GRANOS DE POLEN
Araceae
Dieffenbachia sp. f 1 3
Araceae sp1. f 1 1
Araceae sp2. f 1 3
Arecaceae
Arecaceae sp1. f 1 2
Arecaceae sp2. f 1 2
Arecaceae sp3. f 1 2
Asteraceae
Mikania sp. p 2 2 2 3
Asteraceae sp1. p 1 1
Bignoniaceae
cf. Tynanthus sp. p 1 211
cf. Callychlamys sp. p 1 4
Bignoniaceae sp1. p 1 20
Bignoniaceae sp2. p 1 1
Bignoniaceae sp3. p 1 52
Apndice 1. Listado de plantas registradas en las muestras fecales de los murcilagos del valle de Kosipata, indicando sus formas de uso y
las variables frecuencia y abundancia. Frutos (f), nctar-polen (p), indefnido (i), C. brevicauda (Cb), C. perspicillata (Cp), S. erythromos (Se),
S. oporaphilum (So), S. tildae (St), A. caudifer (Ac), nmero de muestras (n), frecuencia (F), abundancia (A).
(contina....)
Muchhala N. & Jarrin-V P. 2002. Flower visitation by bats in cloud
forests of western Ecuador. Biotropica 34(3): 387395.
Muoz-Saba Y., Cadena A. & Rangel-CH J. 1997. Ecologa de
los murcilagos antflos del sector La Curia, Serrania
La Macarena, Colombia. Rev. Acad. Colomb. Cienc.
21(81):473486.
Novoa S., Cadenillas R. & Pacheco V. 2011. Dispersin de semillas
por murcilagos frugvoros en bosques del Parque Na-
cional Cerros de Amotape, Tumbes, Per. Mastozoologa
Neotropical 18(1): 81-93.
Olea-Wagner A., Lorenzo C., Naranjo E., Ortiz D. & Leon-Paniagua
L. 2007. Diversidad de frutos que consumen tres especies
de murcilagos (Chiroptera:Phyllostomidae) en la selva
lacandona, Chiapas, Mxico. Revista Mexicana de Bio-
diversidad 78: 191-200.
Ortega J. & Alarcon I. 2007. Anoura geoffroyi (Chiroptera: Phyllos-
tomidae). Mammalia Species 818: 17.
Pacheco V. & Solari S. 1997. Manual de los Murcilagos Peruanos
con nfasis en las especies hematfagas. Organizacin
Panamericana de la Salud. 74 pp.
Pennington T. D., Reynel C. & Daza A. 2004. Illustrated guide to
the Trees of peru. David Hunt, The Manse, Chapel Lane,
Milborne Port Sherborne, DT9 5DL. England.
Roubik D. W. & Moreno J. E. 1991. Pollen and spores of Barro
Colorado Island. Missouri Botanical Garden (Eds.). USA.
Sanchez-Casas N. & Alvarez T. 2000. Palinofagia de los murcila-
gos del gnero Glossophaga (Mammalia: Chiroptera) en
Mxico. Acta Zool. Mex. (n. s.) 81: 2362.
Sazima M., Buzato S. & Sazima I. 1999. Bat-pollinated fower
assemblages and bat visitors at two Atlantic Forest sites
in Brazil. Annals of Botany 83: 705712.
SENAMHI Servicio Nacional de Meteorologa e Hidrologa del Per.
2010. (en lnea). <http://www.senamhi.gob.pe/main_mapa.
php?t=dHi>. Acceso 02/2010.
Smith N., Mori S. A., Henderson A., Stevenson D. W. & Heald S.
V. 2004. Flowering plants of the Neotropics. New York
Botanical Garden. Princeton University Press. Italia.
Thies W. & Kalko E. K. V. 2004. Phenology of neotropical pepper
plants (Piperaceae) and their association with their main
dispersers, two short-tailed fruit bats, Carollia perspicillata
and C. castanea (Phyllostomidae). Oikos 104: 362376.
165
Dieta de murcilagos filostmidos de Cusco - Per
Rev. peru. biol. 19(2): 159 - 166 (August 2012)
Bombacaceae
Ochroma sp. p 2 6
cf. Matisia sp. p 1 4
Bombacaceae sp1. p 1 1
Bromeliaceae
Pitcairnia paniculata p 1 4 1 5661
Vriesea sp. p 1 3 1 12
cf. Billbergia sp. f 1 2
Bromeliaceae sp1. f, p 2 3
Burseraceae cf. Protium sp. p 1 14
Cecropiaceae Cecropia spp. f, p 2 12 1 7 1 2 3 80 1 4 2 1037
Clusiaceae cf Vismia sp. f 1 1
Cucurbitaceae
Cayaponia sp. p 1 5 3 199
Gurania sp. f 1 1
Cyclanthaceae cf Asplundia sp. f 1 1
Ericaceae Cavendishia sp. f 1 1
Euphorbiaceae
Acalypha cf macrostachya p 1 7
cf Alchronea sp. p 1 5
cf Omphalea sp. f, p 1 2 1 1
cf Sapium sp. f 1 3
Fabaceae
Inga sp. p 1 14
cf Dioclea sp. i 1 1
cf Mucuna sp. p 1 5
Mimosoidea p 1 4
Caesalpinoidea p 1 11
Flacourtiaceae Flacourtiaceae sp1. f, p 1 3 2 5 3 3243
Gentianaceae cf. Chelonanthus sp. p 1 48 2 23
Gesneriaceae Gesneriaceae sp1. f, p 1 3 1 4
Heliconiaceae Heliconia sp. p 1 1 1 6
Malvaceae Abutilon spp. p 2 10 5 4249
Melastomataceae Miconia sp. f, p 1 2 1 3 1 4
Moraceae
Ficus sp. f 1 1
Sorocea sp. i 1 4
Moraceae-
Urticaceae
Moraceae-Urticaceae sp1. i 1 1
Moraceae-Urticaceae sp2. i 1 2
Moraceae-Urticaceae sp3. i 1 1
Moraceae-Urticaceae sp4. i 1 2
Moraceae-Urticaceae sp5. i 1 23
Piperaceae Piper spp. f 4 1615 2 1262 4 1564 2 1450
Rosaceae
Rubus cf urticiifolius f 1 1
Rubus spp. f, p 1 2 1 2 1 9
Rubiaceae
Genipa sp. p 1 3936
Nertera sp. f, p 1 3
Psychotria spp. f, p 1 2 2 2 1 1
cf Warsewiczia sp. p 1 13
Rubiaceae sp1. f, p 1 2 1 1
Rubiaceae sp2. f, p 1 3
Familia Especie Uso
Cb Cp Se So St Ac
F A F A F A F A F A F A
Apndice 1. Continuacin.
(contina....)
166
Maguia
Rev. peru. biol. 19(2): 159 - 166 (Agosto 2012)
Solanaceae
cf. Solanum sp. f 1 1
Solanum-Lycopersicum f, p 1 1 1 2 1 225
Solanaceae sp1. f 1 1
Solanaceae sp2. f 2 3
Ulmaceae
cf Celtis sp. i 1 1 1 1
Trema micrantha i 1 2
cf. Bombacaceae p 1 3840
cf. Fabaceae p 1 58
cf. Solanaceae f 1 9
cf. Ulmaceae i 1 1
Dicotiledonea i 1 1
Indeterminada 1 i 1 7
Indeterminada 2 i 1 1
Indeterminada 3 i 1 1
Indeterminada 4 i 1 1
Indeterminada 5 i 1 1
Indeterminada 6 i 1 1
Indeterminada 7 i 1 1
Indeterminada 8 i 1 1
Indeterminada 9 i 1 1
Indeterminada 10 i 1 1 1 1
Familia Especie Uso
Cb Cp Se So St Ac
F A F A F A F A F A F A
Apndice 1. Continuacin.
167
Reproduccin y alimentacin del cortarrama peruana en El Gramadal
Rev. peru. biol. 19(2): 167 - 173 (August 2012)
Rev. peru. biol. 19(2): 167 - 173 (Agosto 2012)
Facultad de Ciencias Biolgicas UNMSM
ISSN 1561-0837
Reproduccin y alimentacin de Phytotoma raimondii, cortarrama
peruana en El Gramadal, Ancash
Mario Rosina
1
y Mnica Romo
2
Reproduction and food habits and of Phytotoma raimondii, Peruvian
Plantcutter in El Gramadal, Ancash
1 Asociacin Peruana para la
Conservacin de la Naturaleza
APECO, Parque Jos de Acosta
187, Magdalena, Lima 17, Per.
Email Mario Rosina:
kraken54@gmail.com
2 Email Mnica Romo:
romomonica@gmail.com
Presentado: 25/01/2012
Aceptado: 30/09/2012
Publicado online: 09/11/2012
Resumen
Se confrma la existencia de un buen hbitat con buena disponibilidad de alimento y de sitios para reproduccin
de la cortarrama peruana en El Gramadal (Huarmey). Las dos especies ms importantes para su alimentacin
algarrobo (Prosopis pallida) y palo negro (Grabowskia boerhaviifolia), se encuentran en sufciente cantidad
en el lugar. Esta ltima planta tambin es crucial para la reproduccin, ya que todos los nidos hallados fueron
construidos en esta especie. Hasta el momento se han encontrado nueve nidos entre los aos 2010 y 2012
entre diciembre y abril: siete nidos activos, 2 el 2010 (abril), 4 el 2011, (marzo y diciembre) y uno el 2012
(enero). Tambin se encontraron dos nidos vacos el 2011 (marzo y junio), adems de volantones e individuos
juveniles desde diciembre a junio.
Palabras clave: Phytotoma raimondii; Peruvian Plantcutter; Cortarrama peruana; nidos; volantones; Grabowskia
boerhaviifolia; palo negro.
Abstract
The occurrence of good habitat with suitable food and sites for reproduction for the Peruvian plantcutter at El
Gramadal (Huarmey) is confrmed. The two most important species for food algarrobo (Prosopis pallida) and
palo negro (Grabowskia boerhaviifolia) both occur in suffcient amount in this place. The last species is also
crucial for the reproduction since all the nests found were in this species. To date 9 nests have been found during
2010 and 2012 and between December and April: seven active nests, 2 in 2010 (April) and 4 in 2011(March
and December) and 1 in 2012 (January). In addition, two empty nests were found in 2011 (March and June),
and fedglings and juveniles from December to June.
Keywords: Phytotoma raimondii; Peruvian Plantcutter; nest; chicks; fendglings; Grabowskia boerhaviifolia;
palo negro.
Introduccin
La cortarrama peruana Phytotoma raimondii Taczanowski
1883, es un ave endmica y de distribucin restringida a la costa
norte del Per, desde Piura hasta Ancash, hasta los 300 m de al-
titud (Schulenberg et al. 2010) aunque otros autores mencionan
que llega hasta los 550 m (Clements & Shany 2001). Esta especie
est en peligro de extincin debido sobre todo a la prdida de
su hbitat. El lugar ms al sur donde se le ha encontrado en la
actualidad es el sitio conocido como El Gramadal en la provincia
de Huarmey, departamento de Ancash, donde la poblacin sera
de aproximadamente 80 individuos.
Se conoce de sus hbitos alimenticios y reproduccin por
observaciones realizadas en los departamentos de Piura, Lam-
bayeque, La Libertad y ahora en Ancash. En un trabajo sobre su
dieta en Talara (Abramonte 2007) se indica que la cortarrama
peruana se alimenta de 10 especies de plantas, las que fueron
encontradas por observacin directa, lavado del tubo digestivo
o en heces. En el Santuario Histrico Bosque de Pomac se
menciona que se alimenta de canutillo o palo negro (Gra-
bowskia boerhaaviifolia; E. Sanchez com. pers). En este trabajo
reportaremos la dieta observada en El Gramadal y tambin las
similitudes halladas con las observaciones en los sitios anterior-
mente mencionados.
Sobre la reproduccin, Abramonte (2007) cita que sta ocurre
entre enero y marzo en Talara y describe la conducta de cortejo y
cpula as como el proceso de construccin del nido por ambos
padres. En el Gramadal ya se han descrito dos nidos hallados el
2010 (Rosina & Romo 2010), los primeros reportados desde
1934 (Flanagan & Millen 2008).
El objetivo de este trabajo es ampliar el conocimiento de la
historia natural de la cortarrama peruana, especfcamente de la
alimentacin y reproduccin en el sitio El Gramadal.
Material y mtodos
Lugar de estudio.- El trabajo fue realizado en El Gramadal,
distrito de Huarmey, provincia de Huarmey, Departamento de
Ancash. El sitio es aparentemente el lmite sur de la distribucin
de esta ave y no est registrado en la ms reciente publicacin sobre
la distribucin del Phytotoma raimondii (Flanagan et al. 2009).
La mayor densidad poblacional del ave y la mejor zona de
anidacin se encuentra en la porcin Este, llamada desde ahora
El Gramadal 1, siendo la del lado Oeste El Gramadal 2. Hemos
estimado una poblacin de 40 individuos en El Gramadal 1 y
otros 40 individuos en El Gramadal 2 (Fig. 1).
La superfcie de ambos sitios es de aproximadamente 134
ha, teniendo la zona central o ncleo de El Gramadal 1, 17 ha,
y El Gramadal 2, una extensin de 127 ha respectivamente.
Mientras El Gramadal 1 tiene un suelo formado por sedimento
aluvial proveniente de un antiguo cauce de avenida, hoy seco, El
Gramadal 2 tiene un suelo donde predomina la arena acarreada
por el viento desde el litoral y abundan las pequeas dunas, de-
bido a su cercana de la orilla marina. Este trabajo est centrado
principalmente en el sitio que hemos denominado El Gramadal
1 para el caso de las mediciones de nidos.
168
Rosina & Romo
Rev. peru. biol. 19(2): 167 - 173 (Agosto 2012)
El hbitat consta sobre todo de grama salada, arbustos
achaparrados y espinos, encontrndose un total de 11 especies
vegetales. Adems de registrar las especies en el rea hicimos dos
parcelas de 0,5 hectreas cada una en El Gramadal 1 y otras dos
parcelas de iguales dimensiones en El Gramadal 2 para hallar
la densidad de plantas. En cada parcela se contaba el nmero
de individuos de cada especie de arbusto o rbol. No se cont
la Distichlis spicata grama salada, especie que aparentemente
carece de importancia para la presencia y abundancia del ave
(Tabla 1). Adems de las plantas del sitio, tambin anotamos
las especies de aves con las que la cortarrama peruana comparte
el lugar (Tabla 2).
Se visit El Gramadal los das 29 de noviembre y 28 de di-
ciembre del 2009, 2 de enero, 1, 6, 13 y 14 de abril, 7 y 10 de
Figura 1. Paisaje de El Gramadal en Ancash (foto M.Tweddle).
Especie
Nombre
comn
Forma de
vida
Interaccin con la
cortarrama
Estado de conservacin
de DGFF
1
G1 G2 G1
2
G2
Grabowskia boerhaviifolia
palo
negro o
canutillo
arbusto
alimento (hojas
y frutos), nido,
refugio, posadero.
X X 21,24 14,4
Capparis scabrida zapote arbusto
posadero,
alimento? (hojas)
En peligro crtico (CR) X 3,3
Capparis cordata satuyo arbusto posadero, refugio X X 5,9 5,5
Schinus molle molle rbol
posadero, refugio,
material para nido
X 2,2
Prosopis pallida algarrobo
rbol
achaparrado
alimento (fores,
ramitas tiernas y
brotes), posadero,
refugio
Vulnerable (VU) X X 0,1
Acacia macracantha faique
rbol
achaparrado
posadero Casi amenazado (NT) X 8,2
Distichlis spicata
grama
salada
hierba X X
Cryptocarpus pyriformis chope arbusto X X 0,5
Cordia lutea overo arbusto X
Galvezia fruticosa arbusto X
Ficus carica higo arbusto Introducida X
Total 9 7 6 3
Tabla 1. Especies de fora y nmero de individuos de arbustos y rboles en las dos parcelas en El Gramadal 1 y el Gramadal 2.
1= Direccin General Forestal y de Fauna, Ministerio de Agricultura. 2= nmero de individuos en parcelas de 0,5 ha.
169
Reproduccin y alimentacin del cortarrama peruana en El Gramadal
Rev. peru. biol. 19(2): 167 - 173 (August 2012)
octubre del 2010, 7, 8 y 27 de marzo, 16, 17 y 18 de junio, 23
de julio, 1 de agosto, 26 de diciembre del 2011 y 16 de enero
del 2012 totalizando 19 das en 15 visitas.
Alimentacin.- La cortarrama peruana es un ave exclusiva-
mente vegetariana que utiliza su pico de bordes aserrados para
cortar las ramitas, brotes, hojas y frutos de los que se alimenta.
Cada vez que se visit el lugar se busc indicios de alimenta-
cin en las plantas y se observ las que consuma. Para el caso
del algarrobo, se registr la posicin geogrfca de esta especie
vegetal mediante GPS.
Reproduccin.- En cada visita a El Gramadal se busc ex-
haustivamente nidos en los arbustos y/o conductas de la hembra
o el macho que indicaran construccin del nido o incubacin,
as mismo se observ si haba presencia de volantones o indi-
viduos juveniles.
Resultados
Hbitat.- De las 11 especies vegetales presentes en el rea,
la cortarrama peruana tiene alguna interaccin al menos con 6
de ellas ya sea como alimento, lugar de nidifcacin, posadero,
refugio o material para construccin del nido. En la Tabla 1 se
muestran las especies de arbustos y rboles del lugar y el nmero
de individuos encontrados en cada una de las 2 parcelas de 0,5
ha y su relacin con el ave. En algunos casos algunas especies
presentes se encontraban fuera de las parcelas. El algarrobo
(Prosopis pallida, Fabaceae), presente en este sitio con individuos
achaparrados y en forma arbustiva, se encuentra en una de las dos
parcelas con 1 individuo, pero en las 4 ha donde est el ncleo de
El Gramadal 1 hay 4 individuos. En El Gramadal 2 tambin se
encuentran 8 individuos en una superfcie de 4 ha. La segunda
especie vegetal de gran importancia para el ave es el palo negro o
canutillo (Grabowskia boerhaaviifolia, Solanacea), que es el arbusto
predominante en el lugar, mide hasta 3 m de altura, es espinoso y
muy enmaraado y es muy importante para la reproduccin de
la cortarrama peruana, ya que en esta planta se encontraron los
9 nidos. La densidad del palo negro en dos parcelas de media
hectrea es de 21 y 24 individuos respectivamente en El Gramadal
1 y de 4 y 14 individuos en El Gramadal 2. Otras especies presentes
de importancia son Capparis cordata satuyo, Capparis scabrida
zapote, Schinus molle molle y Acacia macracantha huarango.
Alimentacin.- La dieta de la cortarrama peruana parece
variar de acuerdo a la disponibilidad estacional de las plantas de
su hbitat. En El Gramadal, hemos observado que el algarrobo
tiene brotes y se observan ramitas comidas desde mayo a enero
y sera uno de los alimentos ms importantes durante gran parte
del ao tanto para los adultos como para los juveniles. En no-
viembre hemos observado adems fores de algarrobo comidas.
Por otro lado hemos visto al ave alimentndose de los frutos
y hojas del palo negro (Fig. 2 y 3). Esta planta, se encontraba
fructifcando cada vez que visitamos el lugar y siempre hubo
alguna con mayor abundancia de frutos muy maduros, sobre
todo en los meses ms calurosos del ao (noviembre a junio).
En diciembre y enero encontramos heces con semillas de palo
negro. Los juveniles tambin se alimentan de brotes, hojas y
ramitas tiernas de algarrobo, adems de hojas y frutos maduros
de palo negro. La densidad de ambas plantas es alta en El Gra-
madal 1y la distancia entre stas en la zona de estudio es corta,
lo que facilita el vuelo de los volantones de un arbusto a otro.
Familia Especies
Cathartidae 1. Cathartes aura
FalConidae 2. Falco sparverius
Burhinidae 3. Burhinus superciliaris
ColumBidae 4. Zenaida auriculata
Strigidae 5. Athene cunicularia
Caprimulgidae 6. Chordeiles acutipenis
troChilidae 7. Rhodopis vesper
troChilidae 8. Amazilia amazilia
troChilidae 9. Myrtis fanny
troChilidae 10. Thaumastura cora
Furnaridae 11. Geositta peruviana
tyrannidae 12. Pyrocephalus rubinus
tyrannidae 13. Muscigralla brevicauda
tyrannidae 14. Tyranus melancholicus
phytotomidae 15. Phytotoma raimondii
hirundinidae 16. Pygochelidon cyanoleuca
mimidae 17. Mimus longicaudatus
CoereBidae 18. Coereba faveola
thraupidae 19. Conirostrum cinereum
Tabla 2. Lista de aves de El Gramadal
Figura 2. Phytotoma raimondii, Cortarrama peruana comiendo fruto
de palo negro(foto M.Tweddle).
Figura 3. Phytotoma raimondii, Cortarrama peruana comiendo hoja
de palo negro(foto M.Tweddle).
170
Rosina & Romo
Rev. peru. biol. 19(2): 167 - 173 (Agosto 2012)
Nidos.- La construccin del nido y la alimentacin de los
pichones estn a cargo de ambos padres, mientras que la incuba-
cin corresponde exclusivamente a la hembra (Fig. 4). La madre
suele ausentarse del nido aun con los pichones recin nacidos
a veces hasta por 20 minutos. El macho suele percharse en una
rama alta cercana al nido en actitud vigilante, vocalizando cons-
tantemente y no permite la presencia de otras cortarramas en su
territorio, siendo este comportamiento muy marcado durante
la reproduccin. Cuando no se estn reproduciendo los machos
se exhiben menos, as mismo sus reclamos disminuyen en fre-
cuencia e intensidad. No hemos podido observar apareamientos.
Los nidos en forma de plato son amplios y poco profundos,
miden unos 15 cm de dimetro y su estructura y diseo facilita
el pronto abandono de ste por parte de los pichones. Todos
los nidos encontrados estaban construidos sobre palo negro.
Son fabricados con palitos y ramitas secas de palo negro en la
estructura, molle y algarrobo en la superfcie o cama y se disi-
mulan perfectamente con las ramas del arbusto palo negro. Los
nueve nidos hallados estaban a una altura promedio de 1,52
+ 0,30 m y un rango entre 1,20 2,20 m respectivamente,
estando ocho de ellos entre 1,20 y 1,70 m de altura, el nido 7
encontrado el 16 de junio de 2011 estaba a 2,20 m del suelo.
Estas medidas son tambin un refejo de la altura de los arbustos
de palo negro del lugar. En la Tabla 3 se lista la altura de los
nidos, nmero de huevos, pichones y volantones encontrados
en el 2010, 2011 y 2012.
La hembra pone de 2 a 3 huevos de color celeste claro con
manchas marrones en el polo romo (Fig. 5). Segn nuestras
observaciones la incubacin podra durar entre 13 y 19 das. La
descripcin de los primeros huevos y volantones en este lugar
de estudio se encuentra en Rosina y Romo (2010); luego de
estos, hemos encontrado en el 2011 otros seis nidos ms (nidos
3 al 8) y uno el 2012 (nido 9) con similares caractersticas.
Dos de ellos ya estaban inactivos cuando fueron encontrados
(27 marzo, nido 6 y 16 de junio, nido 7). En diciembre del
2011 se encontr un nido con un pichn emplumando y dos
huevos de color beige, que podran ya estar perdidos debido a
la diferencia que tendran si eclosionan con el pichn nacido
varios das antes, el mismo que al momento de ser encontrado
tendra aproximadamente de siete a diez das de edad (Fig. 6).
Volvimos a visitar el nido el 16 de enero de 2012 y en efecto el
pichn ya haba abandonado el nido y poda observrsele como
volantn en un arbusto cercano, mientras que los 2 huevos,
decolorados por el sol an estaban en el nido. Hemos observado
volantones de los nidos 2, 3 y 4 entre marzo y abril del 2011,
adems en tres oportunidades observamos otros volantones sin
haber encontrado el nido correspondiente (El Gramadal 2: 17 de
junio del 2011, 2 volantones; El Gramadal 1: 26 de diciembre
del 2011, 3 volantones y 2 de enero del 2012, 2 volantones).
Se calcula que de los dos nidos encontrados el 2010 se habran
logrado 3 volantones (75% de los huevos). De los nidos 3, 4,
5 y 8 del 2011, que contenan un total de 10 huevos, 6 (60%)
lograron convertirse en volantones. Estos 9 volantones ms los
5 observados sin haber encontrado el nido signifcara que al
menos 14 juveniles se produjeron hasta el momento (de abril
del 2010 a enero del 2012) (Fig. 7).
Los pichones, nidcolas, son de color negro y empiezan a aban-
donar el nido antes de los 20 das de nacidos, aunque no dejan el
arbusto en el que nacieron, movindose y ocultndose dentro de
l (Fig. 8). El palo negro por su estructura enmaraada y espinosa
les brinda una ptima proteccin contra los depredadores. Poco
despus, ya volantones, siguen a los padres por los arbustos cer-
canos al del nido reclamndoles alimento y tambin intentando
alimentarse por s mismos de hojas y frutos de palo negro, estos
ltimos pareceran todava muy duros y difciles de arrancar de su
pednculo cuando no estn maduros. La distancia entre los dos
Figura 5. Phytotoma raimondii, Nido 5 el 8 de marzo de 2011 (foto
M. Tweddle).
Figura 6. Phytotoma raimondii, Nido 8 el 26 de diciembre de 2011
(foto M. Rosina).
Figura 4. Phytotoma raimondii, nido 4, hembra anidando en palo
negro (foto M.Tweddle).
171
Reproduccin y alimentacin del cortarrama peruana en El Gramadal
Rev. peru. biol. 19(2): 167 - 173 (August 2012)
nidos encontrados el 2010 es de 271 m y la distancia ms corta
y ms larga entre los nidos encontrados los primeros meses del
2011, (nidos 3 al 7) es de 21 a 151 m respectivamente, siendo el
promedio de 92,7 + 42,3 m. Es decir la distancia entre los lmites
de cada territorio podra ser de 20 a 150 m.
Siendo el algarrobo una especie importante como alimento
durante los meses siguientes a la salida de los pichones del nido,
se calcul el promedio de distancia entre los nidos y los algarro-
bos ms cercanos. El promedio de la distancia de los 9 nidos al
algarrobo ms cercano es de 45 m y el promedio de distancia de
los nidos a los dos algarrobos ms cercanos es de 52 m. El nido
9 hallado el 16 de enero del 2012 se encontraba en el mismo
arbusto donde estuvo el nido 3 en marzo del 2011, solo que fue
construido mas al interior del arbusto, aunque casi a la misma
altura (1,60 m y 1,70 m respectivamente).
Durante la poca reproductiva se observa que el plumaje de
las hembras y los machos es ms oscuro, sobre todo las listas
marrones de stas. As mismo, el color gris y las marcas de co-
lor rufo de los machos son ms intensos. Es posible que antes
de la reproduccin los adultos muden su plumaje y conforme
transcurre el ao ste se vaya decolorando por la exposicin a la
luz solar y el desgaste natural de las plumas.
Figura 7. Volantn de Phytotoma raimondii, cortarrama peruana (foto
M.Tweddle).
Figura 8. Phytotoma raimondii, Nido 5 el 27 de marzo de 2011(foto
M. Tweddle).
Nido Fecha
Altura
nido (m)
N.
huevos
N.
pichones
N.
volantones
Fue depredado
o fracaso
Observaciones
1 01/04/10 1,20 1
1 07/04/10 1,20 2
1 13/04/10 1,20 1 Se encontr un huevo eclosionado en el nido
2 07/04/10 1,20 2 2 pichones de 10 a 15 das de nacidos.
2 14/04/10 1,20 2 0
3 07/03/11 1,60 2
3 27/03/11 1,60 1 1
4 07/05/11 1,65 2
4 27/03/11 1,65 1 1
Se colect 1 huevo perforado, 1 huevo
eclosion.
5 08/03/11 1,30 3
5 27/03/11 1,30 3 3 pichones de 1 a 2 das de nacidos.
6 27/03/11 1,20 Nido vaco
7 16/06/11 2,20 Nido vaco
8 26/12/11 1,65 2 1 2
2 huevos no eclosionaron y 1 pichn de 7 a 10
das de nacidos.
8 16/01/12 1 1 Volantn en el arbusto.
9 16/01/12 1,70 2 En el mismo arbusto que nido 3
Tabla 3. Caractersticas de los 9 nidos de Phytotoma raimondii encontrados en El Gramadal el 2010, 2011 y 2012.
172
Rosina & Romo
Rev. peru. biol. 19(2): 167 - 173 (Agosto 2012)
Discusin
La informacin de la historia natural de la cortarrama peruana
en aspectos bsicos como su alimentacin y reproduccin est
aumentando con datos de diferentes lugares de su distribucin.
En cuanto al hbitat hemos visto que para el caso de El Gramadal
el ave vive en un matorral desrtico aunque tambin se le ha
reportado en los llamados bosques secos abiertos con sotobos-
que arbustivo y similar composicin de plantas (Schulenberg
et al. 2010).
Hemos observado que en El Gramadal el ave se alimenta de al
menos dos especies vegetales, algarrobo y palo negro. De mayo a
enero estn disponibles los brotes y fores de algarrobo y el resto
de los meses el palo negro tiene frutos maduros y abundantes,
sobre todo de noviembre a junio. Es decir, durante todo el ao
al menos alguna de las dos especies sera la fuente principal de
alimento. Adems hay que considerar que ambas especies son
abundantes o sufcientes en el rea.
Comparando con la lista de Talara de Abramonte (2007),
quien tambin observ que se alimenta principalmente de
algarrobo con una frecuencia del 73% (fores y frutos de enero
a marzo y brotes de abril a junio; no se estudi de julio a di-
ciembre), en El Gramadal, hay otras plantas presentes de las
que podra alimentarse, como fores (pistilos) y brotes tiernos
de zapote Capparis scabrida o brotes de faique Acacia sp. aunque
nosotros no lo hemos observado. Abramonte (2007) tambin
refere que el ave se alimenta de las fores del overo Cordia lutea,
pero esta planta slo se encuentra en El Gramadal 2. Es decir en
ausencia de brotes y fores de algarrobo o frutos de palo negro,
el ave podra alimentarse de estas otras plantas, aunque algunas
estn muy dispersas y son escasas en El Gramadal.
En la Tabla 4, se presenta una lista de las especies de plantas
de las que se conoce que se alimenta. Es interesante que la dieta
de una de las tres especies congneres, P. rutila, en Santa Fe,
Argentina (De la Pea & Pensiero 2003) incluye 9 especies y
entre ellas tambin consume hojas (septiembre) y frutos (junio
a octubre) de Prosopis alba y hojas (octubre a noviembre) de
Grabowskia duplicata. Si lo comparamos con las estaciones en El
Gramadal vemos que durante los meses de invierno P. raimondii
tambin se alimenta de brotes de Prosopis pallida, mientras que
en los meses de verano lo hemos visto alimentndose de frutos
de Grabowskia boerhaaviifolia. Phytotoma rutila en Crdoba,
Argentina (Bucher et al. 2003) consuma Prosopis alba y P. nigra
a lo largo del ao pero con menores valores en verano; de las
4 especies que ms consumia (Lycium cestroides, Prosopis spp.
Celtis tala y Schinus longifolius), Lycium (SOLANACEA) tena
los valores ms altos de protena a lo largo del ao pero Prosopis
spp. alcanzaba valores similares en el invierno y primavera. Para
Prosopis, la hemicelulosa alcanzaba niveles ms altos en el verano
(22%) que en el resto del ao (1,8 a 13%). Un anlisis de regre-
sin muestra que el contenido de fenoles (r=-0,64, P=0,01) y
protenas (r= 0,59, P= 0,03) eran los factores que ms relacin
tenan con la variacin en la composicin de la dieta. Es lgico
que para las aves folvoras haya seleccin por la mayor cantidad
Nombre cientfco * (comn) e f m a m jn jl a n d por Lugar autor
Prosopis pallida
(algarrobo)
f, fr f, fr f, fr br br br H Talara CA
Prosopis julifora br O Talara CA
Prosopis pallida f br br br br f O Huarmey MR
Grabowskia boerhaviifolia
(palo negro)
fr fr fr, h fr, h fr fr O Huarmey MR
Grabowskia boerhaviifolia f,frbr,h f,frbr,h O Vir LP
Acacia huarango/
macracantha (huarango)
br O Talara CA
Capparis crotonoides
(satuyo)
fr,s s s H Talara CA
Capparis scabrida
(zapote)
f f f H Talara CA
Capparis scabrida h? O Huarmey MR
Maytenus octogona
(realengo)
h h h H Talara CA
Capparis ovalifolia
(vichayo)
br O Talara CA
Cordia lutea
(overo)
f f f O Talara CA
Loxopterigium huasango
(hualtaco)
br O Talara CA
Psitacanthus chanduyensis
(suelda con suelda)
f f f O Talara CA
Tabla 4. Alimentacin de Phytotoma raimondii a lo largo de ao en El Gramadal y otros lugares.
* Diaz 1995.
f=for, fr=fruto, br=brote de hoja, h=hojas, s=semillas. H= en heces, O=Observado comiendo. CA= C.Abramonte 2007, LP=L.Pollack 2009, MR= este trabajo.
173
Reproduccin y alimentacin del cortarrama peruana en El Gramadal
Rev. peru. biol. 19(2): 167 - 173 (August 2012)
de protena ya que las hojas en general tienen poco contenido
de stas; ms an, hay una relacin negativa entre el contenido
de fenoles y protenas (Robbins 1981).
Sobre la importancia de El Gramadal para la reproduccin, se
ha visto que P. raimondii logra tener xito aun cuando su hbitat
sea de pequeas dimensiones, siempre y cuando la composicin y
estructura vegetal de este no sea alterada y contenga los recursos
mnimos necesarios. Esta consideracin tiene importancia en
proyectos de reforestacin con algarrobo o para restauracin
ecolgica, en los que tambin se debera considerar otras especies
tiles para la cortarrama peruana como el palo negro. Adems,
informacin sobre la distancia entre plantas de importancia para
el ave a lo largo del ao es relevante en el momento de plantear
el diseo de las parcelas a reforestar o restaurar.
El Gramadal contiene dos plantas que parecen cruciales para
la presencia y reproduccin de la cortarrama, el palo negro o
canutillo que garantizara no solo el alimento si no su xito re-
productivo y el algarrobo que asegurara el alimento tanto a los
adultos como a los individuos jvenes durante la mayor parte
del ao. La presencia y abundancia de estas dos especies vegetales
ha permitido que El Gramadal contenga la mayor densidad de
cortarramas peruanas y sea uno de los mejores sitios para esta
especie de los 16 lugares visitados donde la hemos encontrado
a lo largo de la costa peruana (Romo & Rosina en prep). Este
estudio ilustra la importancia de medir los recursos tiles a una
especie para entender su presencia y abundancia y en algunos
casos su efecto en la territorialidad. En el caso de El Gramadal,
con distancias entre nidos de menos de 100 m vemos que varias
parejas podran estar visitando o compartiendo los mismos alga-
rrobos del lugar ya que se han encontrado 5 nidos relativamente
cercanos al mismo tiempo en el 2011.
Nuestros resultados confrman la gran importancia de conser-
var El Gramadal ya que no solo es un buen hbitat sino tambin
aqu se ha observado una buena reproduccin de la especie por
3 aos consecutivos. Este lugar es adems el lmite sur de la
distribucin del ave y uno de los pocos ejemplos de matorral
desrtico espinoso aun intacto en la costa peruana. De este tipo
de hbitat, slo quedan unos pocos parches, muy pequeos y
muy alejados entre s.
Agradecimientos
Agradecemos a Michael Tweddle por las estupendas fotogra-
fas que acompaan el artculo y que son un testimonio de la
biologa del ave. A nuestros asistentes Pablo Csar Romo y Lady
Amaro. A Rob Williams quien nos inform sobre el lugar de
El Gramadal. A Manuel Plenge quien con la amabilidad que lo
caracteriza nos facilit bibliografa y revis nuestro manuscrito.
A Angel Servat quien nos brind alojamiento y grata hospitali-
dad en su casa. Mnica Romo expresa que esta investigacin ha
sido hecha fuera de sus actividades profesionales en la Agencia
de Cooperacin Internacional de los Estados Unidos -USAID.
Literatura citada
Abramonte C. 2007. Conducta reproductiva y dieta de Phytotoma
raimondii (TACZANOWSKI, 1883) Cortarrama perua-
na en el bosque seco de Talara. Tesis para optar el ttulo
profesional de bilogo. Universidad Nacional de Piura.
Bucher E.A, D.Tamburini, A.Abril & P.Torres. 2003. Folivory in
the white-tipped plantcutter Phytotoma rutila: seasonal
variations in diet composition and quality. Journal of Avian
Biology 34: 211-216, 2003.
Clements J. & N. Shany. 2001. A feld guide to the birds of Peru.
Ibis Publishing Co, Verona Italy.
De la Pea, M & J.Pensiero. 2003. Contribucin de la fora en los
hbitos alimentarios de las aves en un bosque del centro
de la provincia de Santa Fe, Argentina. Ornitologa Neo-
tropical 14: 499-513, 2003.
Diaz, A. 1995. Los Algarrobos. Concytec. 207 pp.
Flanagan J.N.M. & B. Millen 2008. First nest and eggs records of
Peruvian Plantcutter, Phytotoma raimondii by O.D.Boggs
Bull. B.O.C. 128(4):271.
Flanagan J.N.M, G. Engblom, I. Franke, T. Valqui & F. Angulo
2009. Distribution of the Peruvian Plantcutter Phytoto-
ma raimondii (Passeriformes: Cotingidae). Rev.per.biol.
16(2): 175-182.
Robbins, C. T. 1981. Estimation of the relative protein cost of re-
production in birds. Condor 83:177-179
Rosina M. & M. Romo. 2010. Hallazgo de dos nidos activos de
Phytotoma raimondii, Taczanowski, 1883, cortarrama
peruana. Rev. Per. Biol 17(2): 257-259.
Schulenberg T.S, D.F. Stotz, D.E. Lane, J.P. ONeill & T.A. Parker
III. 2010. Aves de Per. Serie Biodiversidad Corbidi 01.
Lima, Per.
174
Rosina & Romo
Rev. peru. biol. 19(2): 167 - 173 (Agosto 2012)
175
Tallas de lepiDochelyS olivacea en Tumbes, Per
Rev. peru. biol. 19(2): 175 - 180 (August 2012)
Rev. peru. biol. 19(2): 175 - 180 (Agosto 2012)
Facultad de Ciencias Biolgicas UNMSM
ISSN 1561-0837
Estructura de tallas de tortuga pico de loro Lepidochelys olivacea
(Testudines: Cheloniidae) en Tumbes, Per
Manuel Vera
1
y Carlos A. Rosales
2
Size structure of Olive Ridley turtle Lepidochelys olivacea (Testudines:
Cheloniidae) in Tumbes, Peru
1 IMARPE, Instituto del Mar del
Per Laboratorio Costero de
Tumbes, Calle Jos Olaya S/N,
C.P. Nueva Esperanza, Zorritos,
Contralmirante Villar, Tumbes.
2 Marinazul S.A., Calle Santa
Gertrudes S/N, Aguas Verdes,
Zarumilla, Tumbes
Email Manuel Vera:
mvera@imarpe.gob.pe
Email Carlos A. Rosales:
crosales@marinazul.com
Presentado: 12/04/2012
Aceptado: 29/09/2012
Publicado online: 10/11/2012
Resumen
En el presente trabajo se analiz la estructura de tallas de ejemplares de tortuga pico de loro Lepidochelys
olivacea, varados y capturados incidentalmente en las playas de Tumbes, Per (3389,5S 80362,48W y
35721,3S 805745,72W), desde noviembre de 2006 a octubre de 2011. El rea de estudio se dividi en
tres zonas contiguas, limitadas por las quebradas ms activas (Bocapn, El Rubio y Pea Negra). Se registra-
ron 39 ejemplares (74,4% varados y 25,6% capturados incidentalmente), cuyas tallas variaron de 45 a 75 cm
LCC (62,55,7 cm LCC). El 64,1% correspondi a individuos sub-adultos (<65 cm LCC; n= 25). Espacialmente
la mayor cantidad de registros ocurrieron en las zonas 3 y 2, en las cuales porcentajes superiores al 60% se
consideraron sub-adultos; mientras que en la zona 1, el 83% fue adulto. Temporalmente en la poca lluviosa
se registr la mayora de las observaciones (59%), en la cual el 78,3% de ejemplares se consider subadulto;
siendo mayor que en la poca seca (43,8%). Las tallas presentaron diferencias signifcativas entre zonas y
pocas climticas. Se corrobor el patrn de distribucin latitudinal por estados de madurez aparente (ms
adultos en el norte). La presencia del componente adulto indicara que el litoral de Tumbes sera una potencial
zona de anidamiento de L. olivacea, con mayores probabilidades de ovoposicin en poca seca, corroborada
por los eventos de anidamiento recientes.
Palabras clave: Tortugas marinas; estados de madurez aparente; zona potencial de anidamiento; conserva-
cin; mar tropical.
Abstract
In this paper we analyzed the size structure of Olive Ridley turtle Lepidochelys olivacea, from individuals stranded
and incidentally caught in beaches of Tumbes, Peru (3389.5S 80362.48W; 35721.3S 805745.72W),
since November 2006 to October 2011. The study area was divided into three contiguous areas, which were
limited by the most active streams (Bocapan, El Rubio and Pea Negra). There were 39 individuals (74.4%
stranded and 25.6% caught incidentally), with sizes ranged from 45 to 75 cm CCL (62.55.7 cm CCL). 64.1%
was considered sub-adult individuals (<65 cm CCL, n= 25). Spatially the largest number of records occurred
in zones 3 and 2, in which percentages above 60% were considered sub-adults, while in zone 1, 83% was
adult. Temporarily in the rainy season were registered the majority of the observations (59%), in which 78.3%
of specimens was sub-adult, being higher than in the dry season (43.8%). Sizes showed signifcant differences
between zones and climatic seasons. It was confrmed the latitudinal distribution pattern by apparent maturity
stages (more adults in the north). Adults occurrences suggest that Tumbes coast would be a potential nesting
area of L. olivacea, most likely to oviposit in dry season, it which is supported by recent nesting events.
Keywords: Marine turtles; apparent maturity stages; conservation; potential nesting area; tropical sea.
Introduccin
La tortuga golfna o tortuga pico de loro, Lepidochelys olivacea
(Eschscholtz, 1829), es una de las cinco especies de tortugas
marinas que usan el mar peruano como rea de forrajeo en sus
movimientos migratorios y como hbitat de desarrollo de indivi-
duos jvenes (Hays-Brown & Brown 1982, Rosales et al. 2010).
Es categorizada por la Unin Internacional para la Conservacin
de la Naturaleza como especie vulnerable, pues su poblacin est
disminuyendo debido a la explotacin selectiva (recoleccin de
huevos y captura de adultos), captura incidental, deterioro de
los lugares de anidamiento, enfermedades y depredacin (Abreu-
Grobois & Plotkin 2008).
Lepidochelys olivacea es ms comn en la zona norte del Per,
donde las temperaturas superfciales del mar son ms elevadas
(Hays-Brown & Brown 1982), sin embargo su distribucin
tambin ha sido registrada en aguas costeras y ocenicas (IMAR-
PE 2011), Tumbes (Kelez et al. 2003, Forsberg 2008, Rosales
et al. 2010), los puertos Salaverry, Chimbote, Cerro Azul e
Ilo (Alfaro-Shigueto et al. 2002, 2007, 2008), la provincia de
Pisco (Hays-Brown & Brown 1982, de Paz et al. 2002, 2008,
Manrique et al. 2003, 2004) y el puerto San Juan de Marcona
(Majluf et al. 2002).
En Tumbes, L. olivacea es la segunda especie de tortuga ma-
rina con ms registros de varamiento y capturas incidentales,
despus de Chelonia mydas, siendo ms frecuente en Punta Picos,
Canoas y Baja de Punta Mero (Rosales et al. 2010).
En este trabajo se analiza la estructura de tallas de los va-
ramientos y capturas incidentales de L. olivacea en el litoral
de Tumbes, Per, con la fnalidad de evaluar su condicin
poblacional, determinar el porcentaje de estados de madurez
aparente (especialmente del componente adulto) y comprobar las
diferencias en las tallas por zonas de registro y pocas climticas.
176
Vera & Rosales
Rev. peru. biol. 19(2): 175 - 180 (Agosto 2012)
Material y mtodos
rea de estudio.- El estudio se realiz en el litoral de
Tumbes, entre las playas Barrio El 19, La Cruz (3389,5S
80362,48W), y Punta Sal Chico (35721,3S
805745,72W), desde la lnea de costa hasta una milla nutica
de distancia (Fig. 1). El rea fue dividida en tres zonas, cuyos
lmites fueron las quebradas ms activas: 1 (Barrio El 19 a Que-
brada Bocapn), 2 (Quebrada Bocapn a Quebrada El Rubio)
y 3 (Quebrada El Rubio a Quebrada Pea Negra). En esta rea
se distinguen dos pocas climticas: una de lluvias de diciem-
bre a mayo, y otra seca de junio a noviembre. Presenta algunas
playas amplias que en su mayora son de suave pendiente, con
escasa vegetacin y de fcil acceso desde el mar (Rosales et al.
2010), las que presentan numerosas quebradas que permanecen
activas en pocas de lluvia y secas el resto del ao (Ordinola et
al. 2010). En el mar adyacente se ubican ocho bancos naturales
de ostra Crassostrea iridescens, entre 2 y 7 m de profundidad; los
principales bancos se ubican en Punta Sal Chico, Playa Florida
y Bonanza; la pendiente de la plataforma continental es mayor
en la zona sur (entre Lavejal y Punta Sal), predominando los
sedimentos tipo arenoso y rocoso-arenoso (Ordinola et al. 2008).
Colecta de informacin y muestreo.- La informacin bio-
mtrica de L. olivacea provino de la base de datos de los registros
de varamientos y capturas incidentales de tortugas marinas en
Tumbes, recolectados en los recorridos de playas y exploracio-
nes en el mar efectuados durante noviembre de 2006 a julio de
2008, y fue complementada con los datos recolectados en los
recorridos de playas efectuados en agosto de 2009, octubre de
2010 y durante enero, febrero y octubre de 2011, por personal
cientfco de IMARPE Tumbes.
Los ejemplares encontrados fueron identifcados segn Prit-
chard y Mortimer (2000) y Wyneken (2001), y medidos en su
largo curvo del caparazn nucal-supracaudal (LCC), con una
cinta mtrica fexible (mm) (Bolten 2000).
Anlisis de datos.- Se determinaron los parmetros biomtri-
cos de los ejemplares (mnimo, mximo, promedio y desviacin
estndar) y el porcentaje de los estados de madurez aparente
(EMA: sub-adultos y adultos), considerando la talla promedio
de hembras anidantes en la playa El Valle, Colombia (65 cm
LCC; Barrientos y Ramrez 2008).
Los valores de LCC se compararon entre zonas de registro (1,
2 y 3) y entre pocas climticas (lluviosa y seca) con la prueba
de Kruskal-Wallis, debido a que los datos no cumplieron los
supuestos de normalidad y homogeneidad de varianzas.
Resultados
En Tumbes, durante noviembre de 2006 y octubre de 2011,
se registraron 39 ejemplares de L. olivacea, correspondiendo el
74,4% a varamientos y el 25,6% a capturas incidentales. Los
mayores registros ocurrieron en verano de 2007, primavera de
2010 y primavera de 2006 (Fig. 2) y principalmente en Punta
Picos (28,2%), en la zona 2, y en Baja de Punta Mero (25,6%) y
Canoas (15,4%), en la zona 3 (Tabla 1). Las tallas variaron de 45
a 75 cm LCC, con un promedio de 62,55,7 cm LCC (Tabla 2).
Figura 1. rea de estudio (lnea y rectngulo punteados) de Lepido-
chelys olivacea en Tumbes, durante noviembre de 2006 a octubre
de 2011.
Zona Playa Localidad
Total
n %
1
Barrio El 19 La Cruz 1 16,7
Quebrada Charn La Cruz 1 16,7
Nueva Esperanza Nueva Esperanza 2 33,2
Los Pinos Zorritos 1 16,7
Tres Puntas Tres Puntas 1 16,7
Total 6 100,0
2
Bonanza Bonanza 2 12,5
Punta Picos Acapulco 11 68,7
Lavejal Lavejal 3 18,8
Total 16 100,0
3
Baja Punta Mero Punta Mero 10 58,8
Canoas Cancas 6 35,3
Punta Sal Chico Punta Sal 1 5,9
Total 17 100,0
Tabla 1. Nmero de ejemplares de Lepidochelys olivacea por zonas
de registro en Tumbes, durante noviembre de 2006 a octubre de 2011.
Figura 2. Nmero de ejemplares de Lepidochelys olivacea registra-
dos por estaciones del ao en Tumbes, durante noviembre de 2006
a octubre de 2011.
0
2
4
6
8
10
12
14
Pri Ver Oto Inv Pri Inv Inv Pri Ver Pri
2006 2007 2008 2009 2010 2011
N
e
j
e
m
p
l
a
r
e
s
Estaciones del ao
81,0W 80,5W 80,0W
3,5S Pto. Pizarro
La Cruz
Zorritos
Punta Picos
Punta Mero
Canoas
Punta Sal Chico
E
c
u
a
d
o
r
Piura
Tumbes
0 10
Recorridos en
playa
Exploraciones en
el mar
LEYENDA
Zona 1
Zona 2
Zona 3
4,0S
millas nuticas
177
Tallas de lepiDochelyS olivacea en Tumbes, Per
Rev. peru. biol. 19(2): 175 - 180 (August 2012)
De los ejemplares varados (n= 29, todos muertos), el 48,3%
fueron descuartizados para la comercializacin de su carne, y
en algunas ocasiones de su caparazn; y el restante (51,7%)
se mantuvo intacto, aunque el 53,3% present lesiones en el
crneo, plastrn, extremidades y zona gular, que habran sido
causadas por ataques humanos o por colisin con embarcacio-
nes pesqueras. El 37,9% se consideraron adultos (67,32,3 cm
LCC; n= 11).
En cuanto a los ejemplares capturados incidentalmente (n=
10), el 90,0% fue sacrifcado en playa para la comercializacin
de su carne, y en algunas ocasiones de su caparazn (de estos, el
44,4% fueron capturados vivos). Slo un ejemplar fue hallado
muerto (con signos de ahogamiento) y se mantuvo intacto. El
30,0 % se consideraron adultos (69,84,6 cm LCC; n= 3).
Estructura de tallas segn estados de madurez aparente.-
Del total de ejemplares, el 64,1% se consider sub-adulto; mien-
tras que el 35,9% adulto. Las tallas variaron en los sub-adultos
de 45 a 64 cm LCC (59,54,5 cm LCC; n= 25), y en los adultos
de 65 a 75 cm LCC (67,82,9 cm LCC; n= 14) (Tabla 2).
Estructura de tallas segn zonas de registro.- Lepidochelys
olivacea fue registrada en toda el rea de estudio, siendo ms
abundante en la zona 3, entre Quebrada El Rubio y Quebrada
Estadsticos
Estados de madurez aparente
Sub-adulto Adulto Total
n 25 14 39
Mnimo 45 65 45
Mximo 64 75 75
Promedio 59,5 67,8 62,5
DS 4,5 2,9 5,7
n: nmero de ejemplares, DS: desviacin estndar.
Tabla 2. Estadsticos de tallas (largo curvo del caparazn nucal-supra-
caudal: LCC, cm) de Lepidochelys olivacea por estados de madurez
aparente en Tumbes, durante noviembre de 2006 a octubre de 2011.
Zona Playa n Mn. Mx. Prom. DS
1
Barrio El 19 1 69 - - -
Quebrada Charn 1 72 - - -
Nueva Esperanza 2 62 66 64,0 2,8
Los Pinos 1 68 - - -
Tres Puntas 1 68 - - -
Total 6 62 72 67,5 3,3
2
Bonanza 2 64 69 66,5 3,5
Punta Picos 11 48 75 61,7 7,0
Lavejal 3 62 65 64,0 1,7
Total 16 48 75 62,8 6,1
3
Baja Punta Mero 10 57 65 61,3 2,5
Canoas 6 58 68 61,9 4,0
Punta Sal Chico 1 45 - - -
Total 17 45 68 60,5 5,0
n: nmero de ejemplares; Mn.: LCC mnima; Mx.: LCC mxima; Prom.: promedio; DS: desviacin estndar.
Tabla 3. Estadsticos de tallas (largo curvo del caparazn nucal-supracaudal: LCC, cm) de Lepidochelys olivacea por zonas de registro en
Tumbes, durante noviembre de 2006 a octubre de 2011.
0,0
10,0
20,0
30,0
40,0
50,0
60,0
70,0
80,0
Punta Picos Baja Punta
Mero
Canoas
L
C
C
(
c
m
)
Zonas
Figura 3. Promedio de largo curvo del caparazn nucal-supracaudal
(LCC, cm), y desviacin estndar, de Lepidochelys olivacea por zonas
de registro en Tumbes, durante noviembre de 2006 a octubre de 2011.
Pea Negra (Tabla 1). En la zona 1, las tallas variaron de 62 a
72 cm LCC (67,53,3 cm LCC; n= 6); mientras que en la zona
2, de 48 a 75 cm LCC (62,86,1 cm LCC; n= 16); y en la zona
3, de 45 a 68 cm LCC (60,55,0 cm LCC; n= 17) (Tabla 3).
En la zona 1, el 83,3% de ejemplares se consider adulto
(68,62,2 cm LCC; n= 5); mientras que en las zonas 2 y 3,
el 62,5% (59,64,8 cm LCC; n= 10) y 82,4% (59,44,6 cm
LCC; n= 14) se consideraron sub-adultos. Segn la prueba de
Kruskal-Wallis, las tallas presentaron diferencias signifcativas
entre zonas de registro (Tabla 4).
Las playas con mayores registros (ms de tres ejemplares)
fueron Punta Picos (zona 2), Baja de Punta Mero y Canoas (zona
3), las que presentaron promedios de LCC similares durante el
estudio (Fig. 3), con amplio rango de tallas (Tabla 3).
Estructura de tallas segn pocas climticas.- Del total de
ejemplares, el 59% se registr en la poca lluviosa, y el restante
178
Vera & Rosales
Rev. peru. biol. 19(2): 175 - 180 (Agosto 2012)
en la poca seca. En la poca lluviosa, las tallas variaron de 45
a 69 cm LCC (60,55,6 cm LCC; n= 23); mientras que en la
poca seca, de 58 a 75 cm LCC (65,34,6 cm LCC; n= 16)
(Tabla 4). Los mayores promedios de LCC se registraron en
la poca seca, principalmente en 2009, 2010 y 2011 (Fig. 4).
En la poca lluviosa, el 78,3% de ejemplares se consider
sub-adulto (58,95,1 cm LCC; n= 18); mientras que en la poca
seca, el 56,3% se consider adulto (68,63,3 cm LCC; n= 9)
(Tabla 5). Segn la prueba de Kruskal-Wallis, las tallas presen-
taron diferencias signifcativas entre pocas climticas (Tabla 4).
Discusin
Rosales et al. (2010), afrman que el litoral de Tumbes es una
importante zona de forrajeo y desarrollo de L. olivacea, pues su
presencia se registra casi todos los meses del ao, y recomiendan
realizar un trabajo ms completo para evaluar si corresponde a
un hbitat importante especialmente de ejemplares sub-adultos.
Nuestros resultados sobre la proporcin de individuos adultos
(35,9%), sumado a los anteriores registros de hembras anidantes
y de neonatos (Prez et al. 2001, Kelez et al. 2009, Vera et al.
2010, Forsberg et al. 2012) sealan a Tumbes como la regin
ms importante en Per para L. olivacea.
El hallazgo de un mayor porcentaje de individuos sub-adultos
en nuestras observaciones (64,1%) indicara una posible amenaza
a la estabilidad de las poblaciones, porque son considerados
estadios de vida extremadamente valiosos para la recuperacin
de las mismas (Crouse et al. 1987). Este porcentaje fue ligera-
mente inferior al registrado entre noviembre de 2006 y agosto
de 2009 (75,9%) por Rosales et al. (2010) para la misma rea
de estudio, y por De Paz et al. (2007), en Pisco en el verano de
2005 (72,7%). Al respecto, la captura incidental de L. olivacea
en el Per llega a proporciones elevadas, como las observadas
por De Paz et al. (2004) en el rea de Pisco, entre julio de 1999
y junio de 2000, sobre el componente adulto (83%); mientras
en nuestra misma rea de estudio, Rosales et al. (2010) afrman
que los balsilleros capturaron sub-adultos en porcentajes supe-
riores al 70%, similar a lo registrado en nuestras observaciones.
Kelez et al. (2009), afrman que L. olivacea es una especie
comn en Per y que posee un gran componente adulto en
aguas costeras. Al respecto, el porcentaje de ejemplares adultos
registrado por Rosales et al. (2010), para Tumbes (24,1%), fue
similar a los sealados por De Paz et al. (2007), en Pisco (27,3%),
Variable Dimensin n Prom. DS Rango Rango promedio p-valor
Zona
1 6 67,53,3 62-72 31,50
2 16 62,86,1 48-75 20,66
3 17 60,55,0 45-68 15,32
Total 39 0,011
poca climtica
Lluviosa 23 60,55,6 45-69 16,09
Seca 16 65,34,6 58-75 25,63
Total 39 0,010
n: nmero de ejemplares; Prom.: promedio; DS: desviacin estndar.
poca EMA n Mn. Mx. Prom. DS
Lluviosa
Sub-adulto 18 45 63,5 58,9 5,1
Adulto 5 65 68,5 66,5 1,7
Total 23 45 68,5 60,5 5,6
Seca
Sub-adulto 7 58 64 61,2 1,9
Adulto 9 65 75 68,6 3,3
Total 16 58 75 65,3 4,6
n: nmero de ejemplares; Mn.: LCC mnima; Mx.: LCC mxima; Prom.: promedio; DS: desviacin estndar.
Tabla 4. Prueba de Kruskal-Wallis para comparar el largo curvo del caparazn nucal-supracaudal (LCC, cm) de Lepidochelys olivacea entre
zonas de registro y pocas climticas, con el promedio y rango de tallas, en Tumbes, durante noviembre de 2006 a octubre de 2011.
Tabla 5. Estadsticos de tallas (largo curvo del caparazn nucal-supracaudal: LCC, cm) de Lepidochelys olivacea por pocas climticas y
estados de madurez aparente (EMA) en Tumbes, durante noviembre de 2006 a octubre de 2011.
0,0
10,0
20,0
30,0
40,0
50,0
60,0
70,0
80,0
Lluv. Lluv. Seca Seca Seca Seca Lluv. Seca
2006 2007 2008 2009 2010 2011
L
C
C
(
c
m
)
pocas climticas
Figura 4. Promedio de largo curvo del caparazn nucal-supracaudal
(LCC, cm), y desviacin estndar, de Lepidochelys olivacea por
pocas climticas en Tumbes, durante noviembre de 2006 a octubre
de 2011.
179
Tallas de lepiDochelyS olivacea en Tumbes, Per
Rev. peru. biol. 19(2): 175 - 180 (August 2012)
y por Kelez et al. (2009), en toda la costa peruana (33%). En
general, estos valores fueron ligeramente inferiores a lo registrado
en nuestra investigacin (35,9%), lo cual sumado a su presencia
tanto en pocas lluviosas como secas confrmara que L. olivacea
es una especie comn en la costa peruana.
De los ejemplares registrados en la zona 1 de la regin, el
83% se consider adulto (68,62,2 cm LCC; n= 5). Por el
contrario, en las zonas 2 y 3, porcentajes superiores al 60% se
consideraron sub-adultos (59,64,8 cm LCC; n= 10, y 59,44,6
cm LCC; n= 14; respectivamente). Esto corroborara el patrn
de distribucin latitudinal por estados de madurez aparente
sealado por Rosales et al. (2010).
Los elevados porcentajes de ejemplares sub-adultos registrados
en las zonas 2 y 3 (entre Bonanza y Punta Sal Chico), donde
se ubican los principales bancos naturales de ostra Crassostrea
iridescens (Ordinola et al. 2008), indicaran que esta zona es
adecuada para la alimentacin de la especie, debido a la mayor
pendiente de la plataforma continental (especialmente entre
Lavejal y Punta Sal) y al predominio de sedimentos tipo arenoso
y rocoso-arenoso (Ordinola et al. 2008).
La talla promedio registrada en esta investigacin (62,55,7
cm LCC; n= 39), similar a la sealada por Rosales et al. (2010)
(61,94,9 cm LCC; n= 29), evidenci el patrn de distribucin
latitudinal referido por estos investigadores, debido a que fue
ligeramente superior a la registrada en la zona sur del Per por
De Paz et al. (2004) (60,9 cm LCC).
A diferencia de lo sealado por Rosales et al. (2010), la
comparacin de LCC entre zonas de registro y pocas climticas
mostr diferencias signifcativas en las tallas. Al respecto, en la
presente investigacin, la delimitacin de las zonas de registro
(1, 2 y 3) abarcaron una distribucin geogrfca mayor, lo que
pudo determinar las diferencias sealadas. En cuanto a las po-
cas climticas, las diferencias encontradas pueden deberse a los
mayores registros ocurridos en la poca seca por el incremento
de muestreos sistemticos que habran asegurado la recoleccin
constante de muestras.
Las diferencias en tallas segn pocas climticas indicaran
que habra mayores probabilidades de ovoposicin en poca
seca, pues el 56,3% de los ejemplares registrados en esta poca
se consideraron en estado adulto. Las fechas de anidamiento en
Per sealadas por Kelez et al. (2009), agosto de 2007, y Fors-
berg et al. (2012), octubre de 2011, confrman esta asuncin.
La presencia de ejemplares adultos, especialmente en la zona
1 (entre Barrio El 19 y Quebrada Bocapn), indicara que el
litoral de Tumbes es una zona potencial de anidamiento de L.
olivacea, condicin sugerida por las actividades de anidamiento
recientes de esta especie en Per (Prez et al. 2001, Kelez et al.
2009, Vera et al. 2010, Forsberg et al. 2012) y Ecuador (Alava
et al. 2007). Al respecto, Kelez et al. (2009) mencionan que
estas actividades pueden ser una re-colonizacin de las playas al
lmite sur de su distribucin reproductiva en el Pacfco Orien-
tal, lo que facilitara la colonizacin de playas distantes de las
playas natales respectivas. Sin embargo se recomienda que esta
presuncin sea comprobada realizando entrevistas a pobladores
antiguos de la regin.
Segn Marcovaldi y Laurent (1996, en Martnez & Pez
2000), los sitios de anidamiento de esta especie se caracterizan
por ser playas amplias y de fcil acceso desde el mar, pudiendo
realizarse en plataformas bordeadas por quebradas, donde la
franja de arena es ms amplia (Amorocho 1993, en Martnez
& Pez 2000), o cerca a la desembocadura de ros y estuarios
(Cornelius 1976, en Martnez & Pez 2000). En Tumbes,
las zonas de quebradas que discurren en pocas lluviosas y
son observadas en muchos sectores del rea de estudio (p.e.
Quebrada Charn, Quebrada Bocapn, Quebrada El Rubio,
Quebrada Pea Negra), pueden brindar condiciones adecua-
das para el anidamiento de esta especie, pues poseen amplias
franjas de arena.
Por lo tanto, de acuerdo a nuestras observaciones se pue-
de afrmar que el litoral de Tumbes es una zona potencial de
anidamiento de L. olivacea, con mayores probabilidades de
ovoposicin en poca seca. En este sentido, y de acuerdo a las
recomendaciones de Bjorndal (2000), se deben iniciar los es-
tudios que evalen los mecanismos regulatorios que controlan
los parmetros demogrfcos, como los estudios de nutricin,
hormonas, gentica, fsiologa, enfermedades y comportamiento
en la regulacin de la productividad de las poblaciones (creci-
miento y reproduccin) y sobre todo de mortalidad, para saber
cules son sus fuentes y cul es su nivel de impacto.
Agradecimientos
Al Dr. Jorge Llanos, por las facilidades brindadas para el
registro de datos, a los Blgos. Elky Torres y Edward Morales, a
los Ings. Elmer Ordinola, Solange Alemn y Johnny Robles, y
al Tc. Manuel Vlchez, por su apoyo para el registro de datos,
al Blgo. Percy Montero por su apoyo en la ubicacin geogrfca
de las playas de muestreo, y a la Blga. Nelly de Paz, por su apoyo
en la revisin, crticas y recomendaciones en la elaboracin del
presente trabajo.
Literatura citada
Abreu-Grobois A. & P. Plotkin. 2008. (en lnea). Lepidochelys
olivacea. In: IUCN 2011. IUCN Red List of Threatened
Species. Version 2011.2. <www.iucnredlist.org>. Acceso
17/11/2011.
Alava J.J., P.C.H. Pritchard, J. Wyneken & H. Valverde. 2007. First
documented record of nesting by olive ridley turtle (Le-
pidochelys olivacea) in Ecuador. Chelonian Conservation
& Biology 6:282-285.
Alfaro-Shigueto J., M.F. van Bressem, D. Montes & K. Onton. 2002.
Turtle mortality in fsheries off the coast of Peru. In: A.
Mosier, A. Foley and B. Brost, eds., Proceedings of the
Twentieth Annual Symposium on Sea Turtle Biology and
Conservation. NOAA Tech. Memo. NMFS-SEFSC-477.
p. 91-93.
Alfaro-Shigueto J., P.H. Dutton, M.V. Bressem & J.C. Mangel. 2007.
Interactions between Leatherback turtles and Peruvian
artisanal fsheries. Chelonian Conservation and Biology.
6(1):129134.
Alfaro-Shigueto J., J.C. Mangel, P. Pajuelo, et al. 2008. Bycatch in
Peruvian artisanal fsheries: gillnets versus longlines. In:
A.F. Rees, M. Frick, A. Panagopoulou and K. Williams,
eds., Proceedings of the Twenty-Seventh Annual Sym-
posium on Sea Turtle Biology and Conservation. NOAA
Technical Memorandum NMFS-SEFSC-569. p. 192.
Barrientos K. & C. Ramrez. 2008. Estado actual de Lepidochelys
olivacea en el Valle, Pacfco Chocoano, Colombia. In: S.
Kelez, F. van Oordt, N. de Paz and K. Forsberg, eds. Libro
de Resmenes. II Simposio de Tortugas Marinas en el
Pacifco Sur Oriental. p. 17-21. <http://www.ecOceanica.
org/publicaciones> Acceso 13/05/2010.
180
Vera & Rosales
Rev. peru. biol. 19(2): 175 - 180 (Agosto 2012)
Bjorndal K.A. 2000. Prioridades para la investigacin en hbitats de
alimentacin. In: K.L. Eckert, K.A. Bjorndal, F.A. Abreu-
Grobois and M. Donnelly, eds. Tcnicas de Investigacin
y Manejo para la Conservacin de las Tortugas Marinas.
2000 (Traduccin al espaol). IUCN/CSE Grupo Espe-
cialista en Tortugas Marinas Publicacin N 4. p. 13-15.
Bolten A.B. 2000. Tcnicas para la medicin de tortugas marinas. In:
K.L. Eckert, K.A. Bjorndal, F.A. Abreu-Grobois and M.
Donnelly, eds. Tcnicas de Investigacin y Manejo para la
Conservacin de las Tortugas Marinas. 2000 (Traduccin
al espaol). IUCN/CSE Grupo Especialista en Tortugas
Marinas Publicacin N 4. p. 126-131.
Crouse D.T., L.B. Crowder & H. Caswell. 1987. A stage-based po-
pulation model for loggerhead sea turtles and implications
for conservation. Ecology. 68(5):1412-1423.
De Paz N., J.C. Reyes & M. Echegaray. 2002. Datos sobre captura,
comercio y biologa de tortugas marinas en el rea de Pisco
- Paracas. In: J. Mendo and M. Wolff, eds., I Jornada Cien-
tfca "Bases ecolgicas y socioeconmicas para el manejo
de los recursos vivos de la Reserva Nacional de Paracas".
Universidad Nacional Agraria La Molina. p. 125-129.
De Paz N., J.C. Reyes & M. Echegaray. 2004. Capture and trade
of marine turtles at San Andres, southern Peru. In: M.S.
Coyne and R.D. Clark, eds. Procedings of the Twenty-First
Annual Symposium on Sea Turtle Biology and Conserva-
tion. NOAA Technical Memorandum NMFS-SEFSC-528.
p. 52-54.
De Paz N., J.C. Reyes, M. Echegaray, et al. 2007. Identifcacin y
manejo de hbitats crticos de tortugas marinas en Per:
Paracas, estudio de caso. In: C. Guerra-Correa, A. Falla-
brino, P. Bolados-Daz and C. Turner, eds. VII Simposio
Sobre Medio Ambiente. Estado Actual y Perspectivas de
la Investigacin y Conservacin de las Tortugas Marinas
en las Costas del Pacfco Sur Oriental. Antofagasta, Chile.
p. 28. <http://www.uantof.cl/CREA/simpo%20tortugas.
pdf> Acceso 13/05/2010.
De Paz N., J. Quiones & J. Zeballos. 2008. Anlisis preliminar
de la captura de las tortugas marinas en el Puerto de San
Andrs - Pisco, Per. In: S. Kelez, F. van Oordt, N. de Paz
and K. Forsberg, eds., Libro de Resmenes. II Simposio
de Tortugas Marinas en el Pacfco Sur Oriental. p. 70.
Forsberg K. 2008. Proyecto tortugas marinas: iniciativas y esfuerzos
para la conservacin de las tortugas marinas en Tumbes.
In: S. Kelez, F. van Oordt, N. de Paz and K. Forsberg, eds.,
Libro de Resmenes. II Simposio de Tortugas Marinas en
el Pacfco Sur Oriental. p. 69-70.
Forsberg K., A. Petit & M. Arangena. 2012. Avances en el estudio y
monitoreo de anidacin de tortugas marinas en el norte del
Per. In: A. Indacochea, ed. Libro de Resmenes Digital.
III Congreso de Ciencias del Mar del Per. p. 72. <https://
dl.dropbox.com/u/84039404/Aquahoy/LIBRO%20
DE%20RESUMENES%20III%20CONCIMAR%202012.
pdf> Acceso 16/08/2012.
Hays-Brown C. & W.M. Brown. 1982. Status of sea turtles in
Southeastern Pacifc: Emphasis on Peru. In: K.A. Bjorndal,
ed. Biology and Conservation of Sea Turtles. Smithsonian
Press, Washington D.C. p. 235-240.
IMARPE. 2011. Informe nacional sobre la conservacin de las
tortugas marinas en el Per. Comisin Permanente del
Pacfco Sur (CPPS). Plan de Accin para la Proteccin
del Medio Marino y reas Costeras del Pacfco Sudeste.
Lima, Per. 72 pp.
Kelez S., X. Velez-Zuazo & C. Manrique. 2003. Current status of
sea turtles along the northern coast of Peru: preliminary
results. In: J.A. Seminoff, ed., Proceedings of the Twenty-
Second Annual Symposium on Sea Turtle Biology and
Conservation. NOAA Technical Memorandum NMFS-
SEFSC-503. p. 264-265.
Kelez S., X. Velez-Zuazo, F. Angulo & C. Manrique. 2009. (en
lnea). Olive ridley Lepidochelys olivacea nesting in Peru:
The southernmost records in the Eastern Pacifc. Marine
Turtle Newsletter 126:5-9. <http://www.seaturtle.org/mtn/
archives/mtn126/mtn126p5.shtml> Acceso 05/04/2010.
Majluf P., E.A. Babcock, J.C. Riveros, et al. 2002. Catch and by-
catch of sea birds and marine mammals in the small-scale
fshery of Punta San Juan, Peru. Conservation Biology
16:1333-1343.
Manrique C., S. Kelez & X. Velez-Zuazo. 2003. Hatchlings in Peru:
the frst headstarting experience, In: J.A. Seminoff, ed.,
Proceedings of the Twenty-Second Annual Symposium on
Sea Turtle Biology and Conservation. NOAA Technical
Memorandum NMFS-SEFSC-503. p. 99.
Manrique C., S. Kelez, X. Velez-Zuazo & M. Williams de Castro.
2004. New morphometric data on captured sea turtles at
San Andres Inlet, Pisco, Peru. In: M.S. Coyne and R.D.
Clark, eds., Proceedings of the Twenty-First Annual Sym-
posium on Sea Turtle Biology and Conservation. NOAA
Technical Memorandum NMFS-SEFSC-528. p. 122-124.
Martnez L.M. & V.P. Pez. 2000. Ecologa de anidacin de la tortuga
golfna (Lepidochelys olivacea) en la Playa de La Cuevita,
Costa Pacfca Chocoana, Colombia, en 1998. Actual Biol
22 (73):131-143.
Ordinola E., E. Lpez, I. Gonzales, et al. 2010. Identifcacin y
delimitacin de bancos naturales de invertebrados ma-
rinos, zonas de pesca artesanal y reas propuestas para
maricultura en el litoral de la Regin Tumbes. Inf. Interno
Inst. Mar Per. 83 pp. <http://www.imarpe.gob.pe/tumbes/
noticias/Ident_delimit_B_N_Z_12-04-2010.pdf> Acceso
09/12/2010.
Prez O., A. Lujn & Z. Carrin. 2001. Crianza durante nueve meses
en cautiverio de neonatos de Chelonia mydas tortuga ma-
rina verde en el Centro de Acuicultura La Tuna Carranza
del FONDEPES - Tumbes. 9 pp.
Pritchard P.C.H. & J.A. Mortimer. 2000. Taxonoma, morfologa
externa e identifcacin de las especies. In: K.L Eckert,
K.A. Bjorndal, F.A. Abreu-Grobois and M. Donnelly, eds.
Tcnicas de Investigacin y Manejo para la Conservacin
de las Tortugas Marinas. 2000 (Traduccin al espaol).
IUCN/CSE Grupo Especialista en Tortugas Marinas Pu-
blicacin N 4. p. 23-41.
Rosales C.A., M. Vera & J. Llanos. 2010. Varamientos y captura in-
cidental de tortugas marinas en el litoral de Tumbes, Per,
Rev. peru. biol. 17(3): 293-301. <http://sisbib.unmsm.
edu.pe/BVRevistas/biologia/v17n3/pdf/a04v17n3.pdf>
Acceso 02/09/2011.
Vera M., J. Llanos, E. Torres, et al. 2010. Notas sobre neonatos de
Lepidochelys olivacea (Testudines: Cheloniidae) en pla-
ya Nueva Esperanza, Tumbes, Per. Inf. Inst. Mar Per.
37(3-4): 161-166.
Wyneken J. 2001. The anatomy of sea turtles. US Department of
Commerce, NOAA Tec. Memor. NMFS-SEFSC-470,
172 pp.
181
Patgenos introducidos al Per en post larvas de litopenaeuS vannamei
Rev. peru. biol. 19(2): 181 - 186 (August 2012)
Rev. peru. biol. 19(2): 181 - 186 (Agosto 2012)
Facultad de Ciencias Biolgicas UNMSM
ISSN 1561-0837
Patgenos introducidos al Per en post larvas de Litopenaeus vannamei
importadas
Mervin Guevara y Rubn Alfaro
Pathogen introduced to Peru by trade in postlarvae of Litopenaeus vannamei
Laboratorio de Sanidad Acucola,
Instituto del Mar del Per (IMAR-
PE), Sede Tumbes. Panamericana
Norte Km. 1249, Zorritos, Tumbes,
Per.
E-mail Mervin Guevara:
mguevara@imarpe.gob.pe
E-mail Rubn Alfaro:
ralfaro@imarpe.gob.pe
Presentado: 23/01/2012
Aceptado: 04/08/2012
Publicado online: 10/11/2012
Resumen
El comercio internacional de animales acuticos es uno de los factores ms importantes en la introduccin
involuntaria de patgenos a una regin libre de estos. Los patgenos introducidos han sido causa de enormes
prdidas econmicas en organismos bajo cultivo y daos ecolgicos en poblaciones silvestres. En este trabajo,
utilizando la tcnica de la PCR se analizaron postlarvas de Litopenaeus vannamei importadas desde Ecuador
para ser utilizada en los cultivos de langostinos en Tumbes. La fnalidad fue detectar patgenos como el virus
de la mancha blanca (WSV), virus de la cabeza amarilla (YHV), virus de la necrosis hipodrmica y hemato-
poytica infecciosa (IHHNV), Baculovirus penaei (BP), virus del sndrome de Taura (TSV) y la bacteria de la
hepatopancreatitis necrotizante (NHPB). Se analizaron 177 muestras de postlarvas en el ao 2009 y 274 en el
2010. En el ao 2009 se encontraron cuatro patgenos infecciosos en las postlarvas analizadas, siendo ms
frecuente el IHHNV (9,60%). En el ao 2010 la frecuencia de aparicin de postlarvas infectadas con IHHNV
fue mayor (16,79%). Tanto en el 2009 como en el 2010 se encontraron postlarvas positivas a NHPB, BP y TSV
Palabras Clave: Frecuencia, Litopenaeus vannamei, patgenos, postlarva.
Abstract
International trade of aquatic animals is the most important source for pathogens introduction in new areas,
beginning cause of huge economic losses in growing animals and ecological damage. In this paper, we analyze
Litopenaeus vannamei postlarvae imported for shrimp farming companies of Tumbes from Ecuador, in order
to detect pathogens such as the white spot virus (WSV), yellow head virus (YHV), infectious hypodermal and
haematopoietic necrosis virus (IHHNV), Baculovirus penaei (BP), Taura syndrome virus (TSV) and necrotizing
hepatopancreatitis bacterium (NHPB). For this purpose, 177 samples of postlarvae in 2009 and 274 in 2010 from
different larviculture laboratories of Ecuador were processed by the technique of PCR. In 2009, four infectious
pathogens were present in postlarvae. In 2010 the frequency of occurrence of IHHNV infected postlarvae was
higher (16.79%). Both in 2009 and 2010 were found positive postlarvae NHPB, BP and TSV.
Keywords: Frequency, Litopenaeus vannamei, pathogens, postlarvae.
Introduccin
De acuerdo con la Organizacin Mundial de Sanidad Animal,
(OIE), durante la dcada pasada, la produccin mundial de es-
pecies acucolas se ha visto afectada por numerosas enfermedades
infecciosas, las que han impactado severamente el desarrollo y
sustentabilidad de la acuicultura convirtindose en una restric-
cin primaria para su desarrollo (OIE 2010).
La causa principal de brotes de enfermedades y epidemias, est
dada por la transferencia de agentes infecciosos va transporte
de organismos vivos (OIE 2009, Lightner & Redman 1994,
Lightner 1996, Tang et al. 2003, Briggs et al. 2005). La actual
tendencia del libre comercio y el deseo de los pases de generar
nuevas alternativas para el desarrollo social y econmico son
factores que pueden incrementar el riesgo sanitario y que de
no ser considerados oportunamente pueden hacer fracasar la
acuicultura.
Entre los factores que deben tomarse en cuenta para el con-
trol del riesgo se incluyen el estado sanitario de las especies que
deseamos introducir, los problemas sanitarios que las aquejan,
el riesgo de transferencia en la zona donde las vamos a introdu-
cir y los problemas sanitarios de la zona receptora que pueden
afectar a la nueva especie (Frelier et al. 1993, Lotz et al. 2002
Nunan et al. 2003).
Algunos pases lderes en el cultivo de especies acuticas, han
desarrollado polticas relativamente precisas para evitar la trans-
misin de enfermedades, estos lineamientos se han agrupado en
el Cdigo Sanitario para los Animales Acuticos y en el Manual
de Pruebas de Diagnstico para los Animales Acuticos de la
OIE. De la misma manera, pases productores a menor escala,
han desarrollado lineamientos sanitarios propios para evitar la
transferencia de enfermedades a su territorio. Sin embargo, pese
al arduo trabajo, estos lineamientos no han sido efcaces en la
prevencin de la diseminacin de patgenos virales exticos
(Jimnez et al. 1997, Nielsen et al. 2005, Briggs et al. 2005).
En Amrica Latina la realidad es muy similar, porque aun
cuando se cuenta con lineamientos y normas sanitarias que re-
gulan la entrada de especies acucolas, en varios casos se ha dado
la diseminacin de muchos patgenos exticos. (Wyban et al.
2004, Lightner 2006, Senapin et al. 2007, Aguado et al. 2008).
En ciertos patgenos, como el virus de la Mancha blanca
(WSV, white spot virus) y la Necrosis hipodrmica y hemato-
poytica infecciosa (IHHNV, Infectious hypodermal and hae-
matopoietic necrosis virus) el mecanismo de transmisin trans-
ovum, es una importante fuente de contaminacin de huevos
(Lo et al. 1997, OIE 2010). As mismo, existe la posibilidad de
transmisin vertical del virus del sndrome de Taura (TSV, Taura
syndrome virus) y virus de la mionecrosis infecciosa (IMNV,
infectious mionecrosis virus) que no han sido confrmadas
experimentalmente (Brock 1997, Lightner 1996, OIE 2009).
En el caso de contagio por Baculovirus (BP, Baculovirus penaei)
182
Guevara & Alfaro
Rev. peru. biol. 19(2): 181 - 186 (Agosto 2012)
de los reproductores a su progenie, se puede producir mediante
contaminacin fecal de huevecillos al eclosionar, por tanto sin
las medidas proflcticas adecuadas en la larvicultura, inevita-
blemente las larvas cultivadas, sern infectadas. (Overstreet et al.
1998). As mismo, est ampliamente documentado que el agua
es un vehculo de transmisin horizontal de agentes patgenos
a los diferentes estadios larvarios de peneidos (OIE 2009).
Existen muchos ejemplos de propagacin de enfermedades de
animales acuticos en el mundo, entre los casos ms relevantes,
se puede reportar como ejemplo lo ocurrido con el TSV; este
patgeno se identifc inicialmente en Ecuador, (pas en el cual
caus gran mortalidad a los langostinos cultivados) se dispers
rpidamente a Per, Colombia, pases de Centroamrica, Es-
tados Unidos, Hawai, Brasil y pases de Asia, probablemente a
travs de la transferencia regional e internacional de post larvas
y reproductores de L. vannamei infectados con el virus (Lightner
1996, Tu et al. 1999, Briggs et al. 2005, OIE 2009).
Otro caso es el ocurrido con el IHHNV; este patgeno tuvo
su origen en Asia y se sospecha que su ingreso al continente
americano fue producto de la importacin de Penaeus monodon
a Hawai y posteriormente diseminado a otros pases de Lati-
noamrica por el movimiento transfronterizo sin certifcacin
sanitaria adecuada (Tang et al. 2003). La particularidad de este
virus de producir infecciones latentes en sus hospederos, le
permite una mayor transmisin de la infeccin a su progenie y
a otras poblaciones susceptibles (Madhavi et al. 2002).
Con la aparicin del WSV en Per en el ao 1999, el sector
gubernamental mediante Decreto Supremo N 009-99-PE,
decide suspender por un periodo de 180 das la importacin de
nauplios, post larvas y reproductores de langostino Litopenaeus
sp. Posteriormente esta suspensin fue levantada bajo la condi-
cin de llevar a cabo un estricto cumplimiento de la certifcacin
sanitaria de organismos acuticos de importacin, para asegurar
que las larvas importadas ingresaban al pas libre de YHV y WSV.
El Laboratorio de Sanidad Acucola del IMARPE-Sede
Tumbes mediante Resolucin Ministerial N 393-2001- PE
es el rgano facultado para realizar los anlisis que certifquen
el estatus sanitario de post larvas y reproductores de los crus-
tceos que ingresan al territorio peruano. Con este certifcado,
PRODUCE a travs de la ofcina del SANIPES levanta un acta
de conformidad con el que se da inicio a la importacin. Este
trmite se realiza con el fn de evitar el ingreso de nuevos pa-
tgenos como YHV o incrementar la incidencia de infecciones
endmicas por WSV en los centros de cultivo de langostinos o
en los ambientes naturales, que pongan en riesgo la produccin
de langostinos y el estatus poblacional de las diferentes especies
de peneidos silvestres presentes en el Per .
En el presente trabajo informamos sobre la frecuencia de los
patgenos IHHNV, NHPB, BP, TSV, YHV y WSV en las post
larvas de Litopenaeus vannamei importados al Per en los aos
2009 y 2010.
Material y mtodos
rea de estudio.- Se analizaron post larvas importadas de L.
vannamei, procedentes de laboratorios de maduracin de Ecua-
dor, ubicados en los poblados de Mar Bravo, Punta Carnero,
Ayangue y San Pablo, de la provincia de Santa Elena.
Muestras de post larvas.- Entre febrero y diciembre del ao
2009 se colectaron al azar 177 muestras y entre enero y diciembre
del ao 2010, 274.
Las post larvas rutinariamente son remitidas al Laboratorio de
Sanidad Acucola del IMARPE - Sede Tumbes, para el anlisis
obligatorio de WSV e YHV (agentes patgenos de notifcacin
obligatoria, segn la OIE) con la fnalidad de obtener el certi-
fcado que indique que estn libres de tales patgenos y poder
ingresarlas al pas.
Extraccin de cidos nucleicos.- Las extracciones de ADN
se realizaron individualmente, utilizando el mtodo estndar
CTAB-DTAB (Gustincich et al. 1999, modifcado para tejidos
de langostinos). Se pes 25 mg de macerado de post larvas,
en un microtubo de 1,5 mL y se le agreg 600 L de solucin
DTAB (dodeciltrimetilamonio bromuro 8%, NaCl 1,5 M, Tris
HCl 100 mM pH 8,8 y EDTA 50 mM). Con la ayuda de un
micromacerador de plstico se homogeneiz el tejido, para luego
incubarlo a 75 C por 5 minutos. Despus de la incubacin, se
agreg 0,7 mL de cloroformo HPLC, se mezcl con un agitador
automtico y se centrifug en una microcentrfuga a 12000
rpm por 5 minuto. El sobrenadante (250 L) fue transferido
a un microtubo de 1,5 mL que contena 100 L de solucin
CTAB (5% cetiltrimetilamonio bromuro, 0,4 M NaCl) y 900
L de agua destilada autoclavada, se mezcl, incub a 75 C
por 5 minutos y centrifug a 12000 rpm por 10 minutos; el
sobrenadante fue descartado y el pellet resuspendido en 150 L
de solucin disolvente (NaCl 1,2 M) se incub a 75 C por 5
minutos y centrifug a 12000 rpm por 5 minutos. La solucin
fue transferida a un nuevo microtubo con 300 L de etanol al
95%, se mezcl y centrifug nuevamente a 12000 rpm por 5
minutos, el pellet fue lavado con etanol al 75%, secado por 10
minutos y resuspendido con bufer TE (10 mM Tris-HCl, 1
mM EDTA pH 8).
Las extracciones de ARN se realizaron con un kit de ex-
traccin de ARN de IQ 2000
siguiendo las indicaciones del
fabricante.
Cebadores especfcos para el anlisis por PCR.- Los ce-
badores usados para la realizacin de la PCR fueron especfcos
para cada patgeno evaluado y estn recomendados tanto en el
manual de la OIE como en artculos cientfcos actuales (Tabla1)
Anlisis por PCR.- El anlisis de las muestras de post larvas
fue realizado mediante la tcnica de la PCR simple, para el
descarte de los agentes patgenos causantes de las enferme-
dades Hepatopancreatitis necrotizante (NHP), Necrosis
hipodrmica y hematopoytica infecciosa (IHHN), Baculo-
virosis tetradrica y Sndrome de Taura (TS), adems de la
enfermedad de las manchas blancas (WSD) y cabeza amarilla
(YHD). Los productos de amplifcacin para cada patgeno
estn detallados en la Tabla 1.
Para la deteccin de NHPB, las reacciones se realizaron en
mezclas de 20 L, que contenan 2,0 L de bufer Taq 10X
(100 mM Tris-HCl pH 8,8, 500 mM KCl y 0,8% de Nonidet
P-40), 1,5 mM de MgCl
2
, 0,125 mM de cada dNTPs, 1,25 U
de Taq ADN polimerasa (Fermentas), 10 pmol de cada cebador
y 2 L de ADN extrado. El perfl de la PCR se realiz en un
termociclador (9800 Fast Termal Cycler, Applied Biosystems,
USA) y const de 35 ciclos de 94 C por 30 s, 58 C por 30 s,
72 C por 1 minuto y una extensin fnal a 72 C por 5 minutos.
183
Patgenos introducidos al Per en post larvas de litopenaeuS vannamei
Rev. peru. biol. 19(2): 181 - 186 (August 2012)
Para la deteccin de IHHNV, se utiliz un volumen de
reaccin fnal de 20 L, constituida por 2,0 L de bufer Taq
10X, 2,0 mM de MgCl
2
, 0,2 mM de cada dNTPs, 1,25 U de
Taq ADN polimerasa , 150 ng de cada cebador y 2 L de ADN
extrado. El programa de la PCR (termociclador 9800 Fast Ter-
mal Cycler, Applied Biosystems, USA) const de un ciclo de 94
C por 5 minuto, 35 ciclos de 95 C por 30 s, 55 C por 30 s,
72 C por 1 minuto y una extensin fnal a 72 C por 5 minuto.
Para la deteccin de BP el anlisis fue realizado en un volumen
de reaccin de 20 L, constituido por 2 L de bufer Taq 10X,
1,5 mM de MgCl
2
, 0,2 mM de cada dNTPs, 1 U de Taq ADN
polimerasa, 5 pmol de cada cebador y 2 L de ADN extrado.
La PCR (termociclador 9800 Fast Termal Cycler, Applied
Biosystems, USA) se realiz con un ciclo de 95 C por 5 min,
30 ciclos de 94 C por 30 s, 58 C por 45 s, 72 C por 45 s y
una extensin fnal a 72 C por 5 min.
Se utiliz nested PCR para deteccin de WSV, con los ceba-
dores especfcos para la primera y segunda reaccin (Tabla1). El
volumen fnal de cada reaccin fue 50 L, constituido por 5 L
de bufer Taq 10X, 1,5 mM de MgCl
2
, 0,2 mM de cada dNTPs,
1 U de Taq ADN polimerasa 10 pmol de cada cebador y 1 L
de ADN extrado. La PCR fue realizada en un termociclador
(Termolyne Amplitron II, USA) y consta un ciclo de 94 C
por 5 min, seguido de 30 ciclos de 94 C por 30 segundos, 52
C por 30 segundos y 72 C por 45 segundos y una extensin
fnal a 72 C por 5 minutos.
En el caso de la deteccin de TSV, se us el kit de deteccin
Concepto Azul, siguiendo las indicaciones del fabricante.
Los productos de amplifcacin fueron verifcados en geles
de agarosa al 1%, teido con bromuro de etidio (0,5 g/mL) y
por electroforesis en bufer TAE 1X (Tabla1).
Anlisis de datos.- Los parmetros que se determinaron
basados en la informacin obtenida fueron:
- Frecuencia anual de muestras positivas por agente patgeno.
- Frecuencia mensual de muestras positivas por agente
patgeno.
Resultados
Frecuencia anual de muestras positivas a diferentes agentes
infecciosos.- En el ao 2009, se obtuvieron muestras positivas
al IHHNV con una frecuencia de 9,60%, para los patgenos
Tabla 1. Relacin de cebadores especfcos para cada patgeno evaluado y productos de amplifcacin de la PCR.
Tabla 2. Frecuencia mensual de muestras de post larvas positivas a diversos patgenos, febrero a diciembre de 2009.
Patgeno Cebador Secuencia Producto Referencia
NHPB
pf-1 5 ACG TTG GAG GTT CGT CCT TCA G 3
441 pb Loy et al. 1996
pr-2 5 TCA CCC CCT TGC TTC TCA TTG T 3
IHHNV
77012F 5 ATC GGT GCA CTA CTC GGA 3
356 pb OIE 2006
77353R 5 TCG TAC TGG CTG TTC ATC 3
BP
BPA 5 GAT CTG CAA GAG GAC AAA CC 3
560 pb OIE 2006
BPB 5 ATC GCT AAG CTC TGG CAT CC 3
WSV
1 rx
146F1 5 ACT ACT AAC TTC AGC CTA TCT AG 3
1447 pb OIE 2006
146R1 5 TAA TGC GGG TGT AAT GTT CTT ACG A 3
2 rx
146F2 5 GTA ACT GCC CCT TCC ATC TCC A 3
941 pb OIE 2006
146R2 5 TAC GGC AGC TGC TGC ACC TTG T 3
Mes-2009
Muestras de
postlarvas
analizadas
Frecuencia (%)
WSV BP NHPB IHHNV YHV TSV
Enero 0 0,00 0,00 0,00 0,00 0,00 0
Febrero 0 0,00 0,00 0,00 0,00 0,00 0
Marzo 26 0,00 0,00 0,00 19,23 0,00 4
Abril 25 0,00 0,00 4,00 12,00 0,00 12
Mayo 13 0,00 0,00 0,00 0,00 0,00 0
Junio 19 0,00 0,00 0,00 15,79 0,00 0
Julio 10 0,00 0,00 0,00 10,00 0,00 0
Agosto 16 0,00 0,00 0,00 6,25 0,00 0
Septiembre 19 0,00 0,00 0,00 0,00 0,00 0
Octubre 17 0,00 0,00 0,00 5,88 0,00 0
Noviembre 15 0,00 6,67 0,00 20,00 0,00 0
Diciembre 17 0,00 0,00 0,00 0,00 0,00 0
Total anual 177 0,00 0,56 0,56 9,60 0,00 2
184
Guevara & Alfaro
Rev. peru. biol. 19(2): 181 - 186 (Agosto 2012)
BP y NHPB fue de 0,56% y del 2% para el TSV. Las mayores
frecuencias de deteccin del IHHNV se produjeron dentro
del periodo febrero-marzo (19,2%) y en el mes de diciembre
(17,6%). Los patgenos BP y NHPB se detectaron en abril y
noviembre respectivamente (Tabla 2).
En el 2010, se obtuvo muestras positivas al IHHNV y BP,
siendo su frecuencia de deteccin 16,79% y 6,20% respectiva-
mente y de 0,36% para el TSV (Fig. 1). Cabe sealar que se
encontraron muestras de post larvas positivas a IHHNV durante
todo el ao, siendo el valor ms alto encontrado de 44,4%
correspondiente al mes de octubre (Tabla 3).
Todas las muestras de post larvas de importacin analizadas en
este periodo (2009 2010) resultaron negativas a WSV e YHV.
Discusin
El Per cuenta con normativa (Decreto Supremo N 020-
2001-PE) destinada a impedir la entrada de organismos acuticos
(crustceos, poliquetos u otros invertebrados) positivos a YHV
y WSV, exigindose el certifcado sanitario de importacin
extendido por laboratorios autorizados, a pesar de esto existe
la introduccin ilegal de post larvas por parte de empresas
informales. En los aos 2007 y 2009 la DIREPRO (Direccin
Regional de la Produccin) detect 2400 y 4000 millares de post
larvas respectivamente, ingresadas de forma ilegal provenientes
de Ecuador; adems la Aduana de Tumbes inform sobre 3000
millares en el 2007, 2000 millares en el 2008, y 1000 millares
de post larvas en el 2009 que iban a ser ingresadas a territorio
peruano y que al no contar con las certifcaciones sanitarias
fueron incineradas.
El Laboratorio de Sanidad Acucola del IMARPE est facul-
tado (Resolucin Ministerial N 393-2001-PE), para certifcar,
en las post larvas de importacin, la ausencia de los patgenos
causantes de la enfermedad de la cabeza amarilla (YHD) y de
la enfermedad de las manchas blancas (WSD). Sin embargo las
empresas informales burlan estas medidas sanitarias obligatorias
poniendo en riesgo a toda la actividad langostinera de Tumbes.
Segn nuestros resultados, se encontr que para el 2010 la
frecuencia de muestras positivas al IHHNV y BP se incremento
con respecto al ao anterior. Teniendo en cuenta que el xito
de las producciones de langostino est condicionado a disponer
de una fuente confable de post larvas y preferentemente pro-
ducidas en laboratorios locales (FAO 2004), son preocupantes
estos resultados que indicaran una amenaza a la industria del
cultivo de L. vannamei en Per.
Al parecer, las empresas langostineras no prestan la impor-
tancia debida al estado sanitario de las post larvas que impor-
tan, debido a factores como el costo adicional de los servicios
de diagnstico, el desconocimiento de las normas vigentes o
porque estn seguros que ciertos patgenos en las post larvas
no involucran mayores prdidas en la produccin fnal del
cultivo. Tal es el caso del IHHNV, agente etiolgico que no es
mortal para L. vannamei y que causa retardo en el crecimiento
y ocasionalmente deformidad externa (rostrum o segmentos
abdominales). La infeccin experimental con IHHNV redu-
ce signifcativamente las mortalidades de los langostinos tras
Figura 1. Frecuencia anual de muestras positivas a diferentes agentes
infecciosos en post larvas de Litopenaeus vannamei importados.
Tabla 3. Frecuencia mensual de muestras de post larvas positivas a diversos patgenos, enero a diciembre de 2010.
0
2
4
6
8
10
12
14
16
18
WSV BP NHPB IHHNV YHV TSV
2009
2010
Mes -2010
Muestras de
postlarvas
analizadas
Frecuencia (%)
WSV BP NHPB IHHNV YHV TSV
Enero 32 0,00 0,00 0,00 18,75 0,00 0
Febrero 14 0,00 7,14 0,00 7,14 0,00 0
Marzo 16 0,00 0,00 0,00 25,00 0,00 0
Abril 21 0,00 4,76 0,00 9,52 0,00 0
Mayo 19 0,00 0,00 0,00 10,53 0,00 0
Junio 39 0,00 0,00 0,00 7,69 0,00 2,56
Julio 29 0,00 6,90 0,00 13,79 0,00 0
Agosto 15 0,00 0,00 0,00 13,33 0,00 0
Septiembre 15 0,00 0,00 0,00 40,00 0,00 0
Octubre 18 0,00 11,11 0,00 44,44 0,00 0
Noviembre 35 0,00 14,29 0,00 14,29 0,00 0
Diciembre 21 0,00 28,57 0,00 14,29 0,00 0
Total anual 274 0,00 6,20 0,00 16,79 0,00 0,36
185
Patgenos introducidos al Per en post larvas de litopenaeuS vannamei
Rev. peru. biol. 19(2): 181 - 186 (August 2012)
la exposicin con WSV, a esto se le denomina interferencia
viral. Segn este concepto la presencia de dos o ms virus en
un hospedero causar la inhibicin de la replicacin de uno de
ellos (Tang et al. 2003, Namikoshi et al. 2004, Witteeveldt et
al. 2004, Bright et al. 2005, Melena et al. 2006); de esta ma-
nera los langostinos positivos al IHHNV no mueren cuando
son atacados por el WSV, debido a la proteccin que brinda el
IHHNV frente a la infeccin por WSV.
Sin embargo, esto no debe justifcar su presencia en post
larvas de importacin, porque el IHNNV presenta una alta tasa
evolutiva y una amplia diversidad gentica en las cepas aisladas
(Robles et al. 2010), existiendo el riesgo latente de aparicin de
cepas ms virulentas, que pueden poner en peligro a los cultivos
de L. vannamei y las poblaciones silvestres de L. stylirostriss,
especie para la cual el IHHNV es mortal.
Frente a los riesgos mencionados es necesario contar con
estrategias que permitan evitar el ingreso al Per de nuevas cepas
virulentas de patgenos, aplicndose recomendaciones precisas,
como las especifcadas por la FAO y la OIE, entre ellas desarrollar
normativas estrictas; sistemas de supervisin y las capacidades
para la aplicacin de control y registro de las importaciones de
organismos vivos y evitar as el contrabando; obligar al registro
de empresas langostineras informales; organizar talleres de
manejo sanitario en enfermedades para difundir informacin;
crear un directorio de los laboratorios que ofrecen organismos
certifcados libres de enfermedades; desarrollar y aplicar las
buenas prcticas para el manejo en langostineras; establecer
laboratorios de referencia nacional de patologas para los proce-
sos de intercalibracin y asegurar la calidad de los diagnsticos;
establecer programas de reproduccin de ciclo cerrado para
producir semilla SPF (libres de patgenos especfcos) y SPR
(resistentes a patgenos especfcos); y fnanciar programas de
investigacin en estos temas.
Literatura citada
Aguado Garca N, Boada M, de Donato M. 2008. Deteccin del virus
del sndrome de Taura (TSV)en Litopenaeus vannamei
(Boone) del occidente de Venezuela. Revista cientfca,
FCV-LUZ. XVIII, (2): 134 141 pp.
Briggs M, Funge-Smith S, Subasinghe R, Phillips M. 2005. Intro-
ducciones y movimiento de dos especies de camarones
en Asia y el Pacfco. FAO Documento Tcnico de Pesca.
476: 86 pp.
Brock J. 1997. Special topic review: Taura syndrome, a disease
important to shrimp farms in the Americas. World J.
Microbiol & Technol. 13: 415-418 pp.FAO 2004. Ma-
nejo sanitario y mantenimiento de la bioseguridad de los
laboratorios de post larvas de camarn blanco (Penaeus
vannamei) en Amrica Latina. Documento Tcnico de
Pesca. No. 450. Roma, FAO. 2004. 66p.
Gustincich S, Manfolett G, Del Sal G, Schneider C, Carnici P. 1991.
A fast method for high quality genomic DNA extraction
from whole human blood. Biotechniques. 11 (3): 298-302.
Jimnez R, Barniol R, Machuca M. 1997. An epizootic of an intra-
cellular bacterium in cultured penaeid shrimp (Crustacea,
Decapoda) in the Gulf of Guayaquil, Ecuador. In: Flegel T,
MacRae I. Diseases in Asian Aquaculture III. 305-311 pp.
Lightner D, Redman R. 1994. An epizootic of Necrotizing Hepato-
pancreatitis in cultured penaeid shrimp (Crustacea: De-
capoda) in northwestern Peru. Aquaculture 122:918 pp.
Lightner D. 1996. A handbook of shrimp pathology and diagnostic
procedures for diseases of cultured penaeid shrimp. World
Aquaculture Society. Baton Rouge. Louisiana. USA. 304 pp.
Lightner D.V. 2006. The penaeid shrimp viral pandemics due to
IHHNV, WSSV, STV and YHV: History in the Americas
and current status. 1995. Proc. Special Session on Shrimp
Farming. Aquaculture 95 San Diego. World Aquatic Soc.
On Line: Http://www.Lib.noaa/gov/japan/aquaculture/
proceedings/reporte32/lightner/corrected.pdf.
Lo C, Ho C, Chen C, Liu K, Chiu Y, Yeh P, Peng S, Hsu H, Liu H,
Chang C, Su M, Wang C, Kou G. 1997. Detection and tis-
sue tropism of white spot syndrome baculovirus (WSBV)
in captured brooders of Penaeus monodon with a special
emphasis on reproductive organs. Diseases of Aquatic
Organisms. 30(1): 53-72 pp.
Lotz J, Soto M. 2002. Model of White Spot Syndrome virus (WSSV)
epidemics in Litopenaeus vannamei. Dis Aquat Org
50:199209 pp.
Madhavi R, Janakiram P, Jayasree L, Murthy P. 2002. Occurrence
of concurrent infections with multiple viruses in Penaeus
monodon from culture ponds of north coastal Andhra
Pradesh. Current Science. 82 (11): 1397-1400 pp.
Melena J. 2005. Estudio de la co-infeccin viral IHHNV-WSSV
en el camarn blanco L. vannamei. Boletn Informativo
N 130. Disponible en http://www.cenaim.espol.edu.ec/
publica ciones/quincenal/bquinc130.pdf.
Melena J, Bayot B, Betancourt I, Amano Y, Panchana F, Alday V, Cal-
dern J, Stern S, Roch Ph y Bonami JR. 2006. Pre-exposure
to infectious hypodermal and haematopoietic necrosis virus
or to inactivated white spot syndrome virus (WSSV) confers
protection against WSSV in Penaeu vannamei (Boone) post-
larvae. Journal of Fish Diseases. 29: 589-600.
Morales V, Cullar J. 2008. Gua tcnica - Patologa e inmunologa
de camarones peneidos. Programa CYTEC Red II-D
Vannamei. Panam. 270 pp.
Namikoshi A, Wu J, Yamashita T, Nishizawa T, Nishioka T, Arimoto
M y Muroga K. 2004. Vaccination trials with Penaeus
japonicus to induce resistance to white spot syndrome
virus. Aquaculture 229: 25-35.
Nielsen L, Sang-oum W, Cheevadhanarak S, Flegel T. 2005. Taura
syndrome virus (TSV) in Thailand and its relationship
to TSV in China and the Americas. Dis Aquat Org. 63:
101-106 pp.
Nunan L, Noble B, Le Groumellec M, and Lightner D. 2003. Expe-
rimental infection of Penaeus vannamei by a rickettsia-like
bacterium (RLB) originating from Penaeus monodon.
Diseases of Aquatic Organisms 54:4348 pp.
Oie. 2009. Manual de pruebas de diagnstico para los animales
acuticos. Disponible en http://www.oie.int/esp/normes/
fmanual/e_SUMMRY.HTM
Oie. 2010. Manual de pruebas de diagnstico para los animales
acuticos. Disponible en http://www.oie.int/esp/normes/
fmanual/e_SUMMRY.HTM
Overstreet R, Stuck K, Krol R, Hawkins W. 1988. Experimental
infections with Baculovirus penaei in the white shrimp
Penaeus vannamei as a bioassay. J. World Aquaculture
Soc. 19: 175-187 pp.
Robles-Sikisaka R, Bohonak AJ, McClenaghan LR Jr, Dhar AK
(2010) Genetic Signature of Rapid IHHNV (Infectious
Hypodermal and Hematopoietic Necrosis Virus) expansion
in wild Penaeus shrimp populations. PLoS ONE 5(7):
e11799. doi:10.1371/journal.pone.0011799
Senapin S, Phewsaiya K, Briggs M, Flegel T. 2007. Outbreaks of in-
fectious myonecrosis virus (IMNV) in Indonesia confrmed
by genome sequencing and use of an alternative RT-PCR
detection method. Aquaculture. 266: 32-38.
Tang K, Poulos B, Wang J, Redman R, Hsiu-Hui S, Lightner D. 2003.
Geographic variations among infectious hypodermal and
hematopoietic necrosis virus (IHHNV) isolates and cha-
racteristics of their infection. Dis Aquat Org. 53: 91-99 pp.
186
Guevara & Alfaro
Rev. peru. biol. 19(2): 181 - 186 (Agosto 2012)
Tang K, Durand S, White B, Redman R, Mohney L, y Lightner D.
2003 Induced resistance to white spot syndrome virus
infection in Penaeus stylirostris through pre-infection with
infectious hypodermal and hematopoietic necrosis vrus a
preliminar study. Aquaculture 216: 19-29.
Tu C, Huang H, Chuang S, Hsu J, Kuo S, Li N, Hsu T, Li M, Lin
S. 1999. Taura syndrome in Pacifc white shrimp Penaeus
vannamei cultured in Taiwan. Dis Aquat Org. 38:159-
161 pp.
Witterveldt J, Vlak J, van Hulten M. 2004. Protection of Penaeus
monodon against white spot syndrome virus using a
WSSV subunit vaccine. Fish and Shellfsh Immunology
16: 571-576.
Wyban J, White B, Lightner D. 2004. TSV Challenges Advance
selective breeding in Pacifc White Shrimp. Global Aqua-
culture Advocate.
187
Biorremediacin en relaves de cianuracin adicionando nutrientes y microorganismos
Rev. peru. biol. 19(2): 187 - 192 (August 2012)
Rev. peru. biol. 19(2): 187 - 192 (Agosto 2012)
Facultad de Ciencias Biolgicas UNMSM
ISSN 1561-0837
Optimizacin de la biorremediacin en relaves de cianuracin
adicionando nutrientes y microorganismos
Jasmin Hurtado y Arturo Berastain
Optimization of cyanidation tailings bioremediation adding nutrients and
microorganisms
Laboratorio de Biotecnologa Am-
biental, Facultad de Ciencias y
Filosofa, Universidad Peruana
Cayetano Heredia, Honorio Delga-
do 932, San Martn de Porres, Lima.
Email Jasmin Hurtado: jasminhurta-
do@yahoo.com
Presentado: 24/01/2012
Aceptado: 07/10/2012
Publicado online: 10/11/2012
Resumen
El objetivo del presente trabajo fue encontrar los parmetros ptimos para el desarrollo de los proceso de
bioremediacion de relaves de cianuracin, mediante experimentos factoriales a nivel de columnas con la
adicin de nutrientes y el empleo de bacterias sulfato reductoras (BSR) para estabilizar iones metlicos me-
diante la formacin de sulfuros y bacterias capaces de biodegradar cianuro (BC). Se aislaron BSR de relave
y se us un inoculo previamente aislado de BC. Se realiz una prueba de adaptacin a relave y una prueba
en columnas para probar: (1) el efecto del empleo de 1 y 10 mM de lactato para incrementar la actividad de
BSR; y (2) adicin de 0,1 y 1 mM de acetato de sodio y cido fosfrico para mejorar la actividad de BC. Con
las mejores condiciones encontradas se realiz una prueba a nivel de columnas donde se adicion un cultivo
mixto (BSR-BC) y una mezcla de lactato y acetato en concentraciones de 1 mM y 10 mM. Se encontr que
los nutrientes incrementaban la reduccin de sulfatos un 48% en promedio y la adicin de inoculo un 42%. La
bioremediacion de cianuro se mantuvo en 12% sin efecto en la adicin de nutrientes o inoculo. Se prob en dos
relaves adicionales y la adicin de lactato de sodio 1 mM permiti la induccin de BSR en 8 das en uno de ellos.
Palabras claves: cianuro; relaves; BSR; sulfuros; biorremediacin.
Abstract
The objective of this project was to fnd optimum parameters for the development of bioremediation process
of cyanidation tailings by addition of nutrients and microorganisms. It was tested the use of sulfate-reducing
bacteria (SRB) to stabilize metal ions through the formation of sulphides and; bacteria able to biodegrade
cyanide (BC). Factorial experiments were conducted at level of columns. SRB from Cyanidation tailings were
isolated and BC proceeded from pregnant solution of cyanidation. BSR and BC were adapted to tailings. A
column test was performed to try: 1. the effect of using 1 and 10 mM lactate to increase the activity of BSR
and 2. Addition of 0,1 and 1 mM sodium acetate and phosphoric acid to improve the activity of BC. In order to
optimize parameters, it was tested in columns, effect of a mixed culture (SRB-BC) and nutrients (1 mM lactate
and Acetate 1mM). Nutrients were found to increase the reduction of sulfate to 48% (average) and the addition
of inoculums to 42%. Bioremediation of cyanide was 12%. It was not shown effect on the addition of nutrients
or inoculums on Cyanide degradation. Two additional tailings were tested and the addition of 1 mM sodium
lactate allowed the induction of BSR in 8 days in one of them.
Keywords: Cyanide; tailings; SRB; sulphides; bioremediation.
Introduccin
Los relaves de cianuracin provienen del empleo de cianuro
en el tratamiento de minerales para la recuperacin de metales
valiosos y presentan dos problemas qumicos por un lado el
contenido de sulfatos de metales diversos y por otro el remanente
de cianuro que queda despus de los procesos de recuperacin
de metales, ambos vinculados a graves daos a la salud humana
y ambiental.
El empleo de microorganismos es una alternativa para descon-
taminar los relaves de cianuracin, muchos de ellos son bacterias
sulfato reductoras (BSR) que producen sulfuro de hidrgeno el
cual reacciona con los sulfatos en solucin produciendo sulfuros
de metal los cuales son estables y pueden precipitar (White &
Gadd, 1996), adems se conocen otros microorganismos con
capacidad de degradar cianuro (BC) (Knowles 1976; Luque-
Almagro 2005; Rivera & Hurtado 2001).
Las bacterias sulfato reductoras estn siendo usadas en em-
presas relacionadas con metales para el tratamiento de aguas
de descarga a nivel industrial (Lawrence & Marchant 2006;
Scheeren et al. 1992) y en ecosistemas de humedales para trata-
miento de aguas cidas (De Bolder 2003). Para la degradacin
del cianuro se han desarrollado varios procesos, pero a la fecha
solo la compaa Homestake Mining ha desarrollado procesos
para la biodegradacin de cianuro a nivel industrial, el ms
antiguo de ellos viene operando desde 1989 (Witlock 1990).
No se conocen estudios que combinen el empleo de culti-
vos mixtos formados por microorganismos que degraden los
cianuros complejos liberando los iones en solucin, y que a la
vez permitan la estabilizacin de estos mediante la formacin
de sulfuros de metal empleando bacterias sulfato reductoras en
relaves de cianuracin.
El empleo de microorganismos en medios defnidos en
procesos de descontaminacin es conocido, sin embargo las
interacciones que suceden en ellos necesitan ms estudios, por
ejemplo el efecto de agregar nutrientes especfcos a los ambientes
mineros; aunque si existen experiencias agregando nutrientes
para tratar compuestos txicos en otras actividades productivas
(Atlas & Bartha 2002, Scherr et al. 2011)
El objetivo del presente trabajo fue obtener informacin rela-
cionada al uso de nutrientes (lactato, acetato y cido fosfrico) y
188
Hurtado & Berastain
Rev. peru. biol. 19(2): 187 - 192 (Agosto 2012)
microrganismos (BSR-BC) que pueden ser utilizados en procesos
de biorremediacin de relaves de cianuracin. En una primera
fase se us un diseo factorial para optimizar de manera inde-
pendiente la reduccin de sulfatos y la degradacin de cianuro
procedente de un relave de cianuracin, y en una segunda fase
se intent optimizar ambos procesos en conjunto. Finalmente,
se evalu en otros dos relaves el mejor resultado obtenido, en la
induccin de la actividad de bacterias sulfato reductoras.
Material y mtodos
Microorganismos.- Las bacterias reductoras de sulfato (BSR)
fueron aisladas de relaves de cianuracin proporcionados por la
Compaa minera Orcopampa (1516'S, 7219'W). El medio
utilizado para el aislamiento fue el medio de Baars Modifcado
(Starkey 1938), el cual tiene la siguiente composicin (en g/L):
KH
2
PO
4
, 0,5; NH
4
Cl, 1,0; MgSO
4
.7H
2
0, 2,0; CaSO
4
, 1,0;
FeSO
4
.7H20, 0,05; lactato de sodio (70%), 3,5; extracto de
levadura, 1,0 y citrato de sodio, 5,0. El pH fue ajustado a 7 con
NaOH (10% w/v). Se sembraron muestras de 1 g de cada relave
en 9 mL de medio de Baars modifcado y se colocaron en cma-
ras de anaerobiosis con sistemas de generacin de anaerobiosis
AnaeroGen de 2,5 L (Oxoid) a 30 C.
Las muestras fueron mantenidas durante 2 semanas y repi-
cadas en medios empleando el mismo sistema de anaerobiosis.
Se monitoreo el crecimiento mediante microscopa y formacin
de sulfuro de ferro (ennegrecimiento de medio).
Las cepas utilizadas para biorremediacin de cianuro (BC)
fueron tres cepas de Pseudomonas sp. las cuales fueron previamen-
te aisladas de una solucin cargada de cianuracin utilizadas en
procesos de biorremediacion de estas soluciones (Freyre 2000),
las cuales fueron mantenidas en medio PGY con la siguiente
composicin (en g/L): peptona, 2,5; glicerol, 1,25; extracto de
levadura, 0,25; agar 15,0. El pH fue ajustado a 9,0.
Muestras de relave de cianuracin.- Tres muestras de rela-
ves de cianuracin fueron proporcionadas por la empresa BH
Consultores entre mayo de 2008 y junio de 2009. La muestra
de relave 1, tena la siguiente composicin (en mg/L): Ag 0,23;
Cu 19,3; S 891; Sb 4; cianuro Total 193; cianuro WAD 0;
cianuro libre 0 y sulfatos 3070. Los relaves 2 y 3 tenan 939 y
4279 mg/L de sulfatos respectivamente.
La muestra 1 fue utilizada para la determinacin de los
parmetros ptimos. Los relaves 2 y 3 fueron ensayados en los
parmetros optimizados.
Anlisis Qumicos.- Los anlisis de sulfatos fueron realizados
empleando el mtodo 4500-SO4
2
C Sulfate. Gravimetric Me-
thod with Ignition of Residue (APHA, AWWA y WEF 1999).
El cianuro total fue determinado por la Tcnica colorimtrica
4500-CN-N Total Cyanide after destilation (APHA, AWWA y
WEF 1999). El anlisis se llev a cabo con espectrofotmetro
Hewlett Packard 6453.
Adaptacin de bacterias a relave 1 a nivel de frascos.- La
adaptacin de bacterias sulfato reductoras al relave 1 se realiz
sembrando las bacterias aisladas en frascos conteniendo 50 mL
de relave, 50 mL de medio Baars y 10 mL de inoculo BSR.
Se prepar controles en las mismas condiciones sin inoculo.
Los frascos fueron mantenidos en cmaras de anaerobiosis con
sistemas de generacin de anaerobiosis AnaeroGen a 20 C
durante 4 semanas. Se realizaron coloraciones y conteo de bac-
terias empleando microscopio de contraste de fases. Asimismo,
se sembr 0,1 mL del cultivo en medio lquido de Baars en
anaerobiosis para verifcar la presencia y actividad de bacterias
sulfato reductoras.
Para adaptar las bacterias biodegradadoras de cianuro al
relave 1, se sembraron frascos conteniendo 50 mL de relave,
50 mL de medio PGY y 10 mL de inoculo BC. Se prepar un
frasco control sin bacterias. Los frascos fueron sellados con pa-
raflm y mantenidos a 20 C durante 4 semanas. Se realizaron
verifcaciones peridicas del crecimiento mediante coloraciones
GRAM y conteos.
Efecto de nutrientes e inoculo en la reduccin de sulfatos
y biodegradacin de cianuro de relave 1 a nivel de columnas.-
Se determin mediante un diseo factorial 2
2
el efecto del nivel
de adicin de nutrientes e inoculo. Se montaron 3 grupos de
columnas. En el primer grupo se estudio el efecto de lactato de
sodio e inoculo BSR en la reduccin de sulfatos. En el segundo
y tercer grupo se determin el impacto de: (1) la adicin de
acetato de sodio e inoculo BC y (2) la adicin de cido fosfrico
e inoculo BC en la degradacin de cianuro.
Para cada grupo, se montaron 8 columnas de vidrio de 235
mm de altura por 30 mm de dimetro las cuales fueron llenadas
completamente con 120 mL de relave.
Para comprobar el efecto en la reduccin de sulfatos, se agreg
de acuerdo al diseo, solucin stock de Lactato de sodio hasta
dar una concentracin fnal de 1 mM y 10 mM y como inoculo
1 mL de cultivo de bacterias sulfato reductoras.
La verifcacin del efecto de acetato de sodio en la biode-
gradacin de cianuro se realiz empleando una solucin stock
hasta llegar a una concentracin fnal de 0,1; 1 mM y 1 mL de
bacterias degradadoras de cianuro.
El efecto del cido fosfrico fue estudiado empleando una
solucin stock hasta una concentracin fnal de 0,1; 1 mM y 1
mL de bacterias degradadoras de cianuro. Se montaron adems
dos columnas control solo con relaves.
Las columnas fueron cubiertas con paraflm y permanecieron
estticas y a 20C durante 4 semanas. Terminadas las 4 semanas,
las soluciones fueron analizadas para cianuro total y sulfatos.
Determinacin de los parmetros ptimos para la bio-
degradacin de cianuro y reduccin de sulfatos en relave 1
a nivel de columnas.- Se empleo un diseo factorial 2
2
para
determinar el efecto del nivel de adicin de acetato de sodio
y lactato de sodio e inoculo mixto (BSR-BC). Se montaron 8
columnas de vidrio de 350 mm de altura por 50 mm de dimetro
las cuales fueron llenadas completamente con 600 mL de relave.
Para comprobar el efecto en la reduccin de sulfatos, se agreg
de acuerdo al diseo, una solucin stock de lactato de sodio y
acetato de sodio hasta dar un volumen fnal de 1 mM y 10 mM
de lactato de sodio y como inoculo 5 mL de cultivo mixto de
bacterias (2,5 mL de bacterias degradadoras de cianuro y 2,5
mL de bacterias reductoras de sulfato). Se montaron adems dos
columnas control sin nutrientes ni inoculo.
Las columnas fueron cubiertas con paraflm y permanecieron
estticas y a 20 C durante 4 semanas. Terminadas las 4 semanas,
las soluciones fueron analizadas para cianuro total y sulfatos.
189
Biorremediacin en relaves de cianuracin adicionando nutrientes y microorganismos
Rev. peru. biol. 19(2): 187 - 192 (August 2012)
Efecto de la adicin de nutrientes en la induccin de
bacterias sulfato reductoras en los relaves 2 y 3.- Se colocaron
en tubos de prueba (por triplicado) 2 g de relave y 10 mL de
solucin con 0,5; 1 y 2 mM de lactato de sodio y otro set con 2,
1, y 0,5 de acetato de sodio. A los tubos se les coloco papeles em-
bebidos en acetato de plomo (Gerardt et al. 1981). Se colocaron
los tubos en una jarra de anaerobiosis a 30 C durante 1 mes. La
induccin de bacterias sulfato reductoras fue evidenciada por la
formacin de sulfuro de plomo en las tiras de acetato de plomo.
Anlisis Estadstico.- Se realizaron anlisis estadsticos
usando SPSS versin 14. Inicialmente se calcularon los valores
estadsticos descriptivos (valores mnimo, mximo, promedio y
desviacin) y se verifco la normalidad de la distribucin de las
mediciones de las concentraciones de cianuro y sulfato con la
prueba de Kolgomorov-Smirnov, presentndose una distribucin
normal. Posteriormente se aplic la prueba de Levene y la de
ANOVA factorial utilizando como variables dependientes las
concentraciones de cianuro y sulfato y como factores, el empleo
de nutrientes y adicin de inoculo. Todas las pruebas se traba-
jaron con un nivel de signifcancia de 0,05.
Resultados
Aislamiento de bacterias sulfato reductoras.- La siem-
bra de relaves en medio liquido de Baars produjo un ligero
ennegrecimiento del medio a los 5 das, el cual se intensifco
gradualmente hasta encontrarse el medio completamente negro
debido a la formacin de sulfuro de hierro, por la produccin
de sulfuro de hidrgeno.
En coloracin Gram se encontraron bacilos Gram negativos.
Crecimiento y actividad de bacterias sulfato reductoras en
frasco con relave 1.- En la prueba fnal se observ la presencia de
bacilos Gram negativos. Asimismo, el cultivo del medio produjo
formacin de sulfuros (color negro), que nos est indicando la
presencia de bacterias sulfato reductoras creciendo con relave 1.
Crecimiento y actividad de bacterias degradadoras de
cianuro en frasco con relave 1.- Se observ en las revisiones
peridicas, presencia de 5 x 10
8
clulas/mL de microorganismos
con morfologas semejantes a las cepas inoculadas. En la muestra
control se pudo encontrar una poblacin de 10
5
clulas/mL. Apa-
rentemente la poblacin natural de la solucin de cianuracin
incrementada por la adicin del medio de cultivo.
Efecto del lactato de sodio y del inoculo en la reduccin
de sulfatos en relave 1 a nivel de columnas.- En la Tabla 1 se
pueden observar los valores estadsticos descriptivos de todas las
pruebas realizadas a nivel de columnas.
La Tabla 2 muestra los resultados de la aplicacin de lactato
e inoculo BSR en la reduccin de sulfatos. El porcentaje pro-
medio de reduccin de sulfato en estas condiciones fue de 34%
(1069 mg/L). El control de columnas de relave sin ninguna
adicin, produjo una disminucin de sulfatos en solucin del
1% (70 mg/L).
El anlisis factorial 2
2
de los resultados presentados en la
Tabla 2, muestra que la variacin de la adicin de Lactato de
sodio 1 mM a 10 mM aumenta la concentracin de sulfato en
solucin en 500 mg en promedio y que la adicin de inoculo
disminuye la concentracin de sulfato en solucin en 100
mg/L.
La reduccin de sulfato se ve incrementada de manera estads-
ticamente signifcativa cuando se emplea 1 mM de lactato de
sodio y cuando se produce una interaccin de lactato e inoculo.
Efecto de nutrientes e inoculo en la degradacin de
cianuro de relave 1 a nivel de columnas.- Del anlisis de los
resultados obtenidos sobre degradacin de cianuro presentados
en la Tabla 3, se muestra que en la primera serie de pruebas
que emplea acetato de sodio, la degradacin aumenta cuando
se emplea 1 mM de acetato de sodio, sin embargo no hay una
diferencia signifcativa entre emplear 0,1 mM y 1 mM. Si hay
una diferencia estadsticamente signifcativa cuando se emplea
inoculo BC y hay interaccin entre acetato e inoculo.
Puebas Concentracion de valores dependientes (mg/L)
Valor Dependiente Factor n Minimo Maximo Media Desv. Tip.
Sulfato
Lactato-BSR 10 1600 3067 2205 528
Optimizacion 10 1390 3067 2122 571
Cianuro
Acetato-BC 10 120 186 150 19
Fosforo-BC 10 126 186 157 23
Optimizacion 10 144 186 159 12
Tabla 1.Valores estadsticos descriptivos de los valores dependientes (concentracin de sulfato y cianuro) en las 3 pruebas de la primera fase
(Efecto) y las 2 pruebas de la segunda fase (Optimizacin).
Matriz de experimentos Plan de Experimentacin
Sulfato mg/L % Reduccin de sulfato
X1 X2 Lactato de sodio mM Inoculo BSR (mL)
- - 1 mM NO 1625 48
+ - 10 mM NO 2400 22
- + 1 mM SI 1876 39
+ + 10 mM SI 2100 33
Control NO NO 3063 1
Tabla 2. Porcentaje de la reduccin de sulfatos en relave 1 a nivel de columnas obtenidos en la primera fase. Los resultados mostrados son
el promedio de pruebas por duplicado.
190
Hurtado & Berastain
Rev. peru. biol. 19(2): 187 - 192 (Agosto 2012)
En el segundo conjunto de pruebas (en las cuales se empleo
cido fosfrico) se obtienen mejores resultados signifcativos
de degradacin de cianuro, cuando se utiliza 1 mM de cido
fosfrico sin inoculo.
Los controles con relave mostraron una disminucin de cia-
nuro total del 8% (15 mg/L). Por lo tanto, el porcentaje prome-
dio de reduccin de cianuro, eliminado el degradado de manera
natural en el control, fue de 20% (38 mg/L) cuando se emplea
acetato, mientras que el porcentaje promedio de reduccin de
cianuro es 14% (27 mg/L) cuando se emplea fsforo. Debido
al incremento de reduccin producida por la adicin de acetato
de sodio, se decidi seguir con las pruebas empleando acetato
de sodio a concentraciones mayores e inoculo BC.
Determinacin de los parmetros ptimos para la biode-
gradacin de cianuro y reduccin de sulfatos de relave 1.- El
promedio de la reduccin de sulfato fue de 34%(1044 mg/L).
Se encontr diferencias signifcativas entre las condiciones em-
pleadas en las columnas en cuanto a la reduccin de sulfato en
solucin (Tabla 4).
El promedio de disminucin de cianuro fue de 12% (23
mg/L). El anlisis del contenido de cianuro total en solucin no
mostr diferencias estadsticamente signifcativas cuando se us
inoculo mixto ni cuando se agreg nutrientes, sin embargo si
hubo una diferencia signifcativa cuando se produjo interaccin
de nutrientes e inoculo.
Se obtuvo 55% (1390 mg/L) de reduccin de sulfato en
solucin cuando se agrego 1 mM de nutrientes e inoculo mixto.
Cuando se agreg 1 mM de nutrientes al relave 1, se encontr
15% menos de sulfato en solucin que cuando se agrego 10 mM
de los nutrientes. Cuando se agreg inoculo mixto a los relaves
se encuentra un 8% ms de sulfato en solucin que cuando no
se agrega inoculo al relave 1.
El factor ms importante fue el agregar nutrientes (lactato y
acetato) y el efecto ms resaltante fue la reduccin de sulfatos,
por lo cual se decidi probar la adicin de lactato de sodio y
acetato de sodio en la induccin de bacterias sulfato reductoras
a diferentes concentraciones en los 2 relaves adicionales.
Efecto de la adicin de nutrientes en la induccin de
bacterias sulfato reductoras en los relaves 2 y 3.- Se trabaj
con 2 relaves de cianuracin de diferentes procedencias. En el
relave 2 no se pudo evidenciar la presencia la actividad de bac-
terias sulfato reductoras en los 30 das que dur la prueba. En
el relave 3, que contaba con un contenido de sulfatos de 4279
mg/L se demostr la presencia de bacterias sulfato reductoras,
al agregarse lactato de sodio (Tabla 5).
Discusin
Las bacterias sulfato reductoras son un grupo muy diverso de
procariotas que usan la reduccin desasimilatoria de sulfato para
la generacin de energa, por lo que nuestro estudio se dirigi
a investigar su actividad.
Matriz de
experimentos
Plan de Experimentacin
CN- Total (mg)
% degradacin
de cianuro
X1 X2
Nutrientes (X1)
Inoculo BC
(X2) (mL)
Acetato de sodio
(mM)
Acido fosfrico
(mM)
- - 0,1 0 No 162 16
+ - 1 0 No 138 28
- + 0,1 0 Si 129 34
+ + 1 0 Si 144 25
- - 0 0,1 No 136 27
+ - 0 1 No 128 33
- + 0 0,1 Si 181 6
+ + 0 1 Si 150 22
Control No No No 178 8
Tabla 3. Porcentaje de degradacin del cianuro en el relave 1 a nivel de columnas obtenidos en la primera fase. Los resultados mostrados
son el promedio de pruebas por duplicado.
Fuente Suma de cuadrados tipo III gl F Signifcacion
Modelo corregido 724375 3 2143 0.000
Interseccion 28505025 1 253096 0.000
Inoculo 61075 1 524 0.000
Nutrientes 451725 1 4010 0.000
Nutrientes*Inoculo 211575 1 1878 0.000
Error 450 4
Total 29229851 8
Total corregida 724825 7
Tabla 4. Sumario del ANOVA de resultados de la optimizacin de factores (lactato, acetato e inoculo mixto) en la concentracin de sulfato (mg/L).
191
Biorremediacin en relaves de cianuracin adicionando nutrientes y microorganismos
Rev. peru. biol. 19(2): 187 - 192 (August 2012)
Las concentraciones empleadas de lactato de sodio y el uso de
acetato de sodio con el objeto de incrementar la reduccin de sul-
fatos, estn basadas en los estndares empleados en los medios de
cultivo para favorecer el crecimiento de microrganismos (Freyre
2000, Starkey 1938), mientras que la concentracin de fosfato
fue la misma utilizada en el trabajo de Withlock (1990). En este
trabajo, se ha confrmado, que existe efecto de estimulacin de
la actividad de microrganismos en relaves de cianuracin, que
es producido por la adicin de lactato principalmente, incluso
en concentraciones menores de las publicadas.
Se conoca que la adicin de 10 mM de acetato y 10 mM de
lactato bioestimulan la poblacin de BSR, produciendo mayores
velocidades de reduccin de sulfuros en suelos contaminados
con uranio y metales pesados (Sitte et al. 2010), tambin que,
en aguas cidas artifciales, el incremento de celulosa produce
un incremento en la actividad de las bacterias sulfato reductoras
(Bechard et al. 1993). En nuestro trabajo la concentracin de 1
mM de acetato, lactato o fosfato produjeron la disminucin de
sulfatos o cianuro, y en la degradacin de cianuro tuvo el mayor
efecto el acetato que el fosfato.
El lactato es un compuesto comnmente usado en el aisla-
miento de BSR. La utilizacin del acetato divide a las bacterias
sulfato reductoras en dos grandes grupos, aquellas que degradan
compuestos orgnicos hasta acetato y las que degradan com-
puestos orgnicos, incluyendo al acetato, hasta CO
2
(Strattan
2010). En las bacterias sulfato reductoras, el lactato sirve como
aceptor de electrones para la recuperacin de metales, el lactato
o acetato como fuente de carbono y el sulfato como aceptor de
electrones (White & Gadd 1996). Como hemos demostrado el
lactato aadido a los relaves incremento la produccin de sulfuro
de hidrgeno, porque las bacterias sulfato reductoras oxidan el
lactato y usan el hidrgeno producido para la reduccin de sul-
furos (Muyzer & Stams 2008). El uso de lactato 1 mM produjo
el mayor efecto llegndose a un 47% de reduccin de sulfatos.
En las pruebas a nivel de columnas se favorece la produccin
de anaerobiosis en zonas o nichos anxicos que permiten la
degradacin de sulfatos. Algunas especies de bacterias sulfato
reductoras pueden resistir bajas concentraciones de oxgeno,
aunque la reduccin de sulfatos es anaerbica (Muyzer & Stams
2008), adems reduccin de sulfatos puede ocurrir en hbitats
oxigenados en los cuales se forman nichos anxicos (Sitte 2010)
El efecto de la adicin de inculos depende de los microor-
ganismos presentes en ellos y de los ambientes involucrados.
La adicin de bacterias sulfato reductoras fue el segundo efecto
signifcativo ms importante incrementando la reduccin de
sulfatos a un 42%. La adicin de un inculo de bacterias
degradadoras de cianuro no tuvo un efecto signifcativo en
la disminucin de la concentracin de cianuro en la solucin
podra deberse a diferentes motivos. Primero, el inculo BC
empleado para la degradacin de cianuro en esta prueba, fue
aislado de una solucin cargada de cianuracin y la aplicacin
de este inoculo BC permiti la disminucin del 88% (584
mg/L) de cianuro total (Freyre 2000); pero estos microrga-
nismos podran no funcionar de manera efciente con relaves
o soluciones de otras minas, ya que la composicin qumica
y mineralgica de cada relave y solucin cargada esta basada
en la composicin del mineral y, sus caractersticas fsicas
(granulometra, porcentaje de slidos, etc.) dependen de los
procesos de concentracin del mineral (Velarde 1992). Se-
gundo, cuando se trabajo solo con un inoculo BC se obtuvo
en promedio 23,5% de disminucin de cianuro, a diferencia
de cuando se empleo un inoculo mixto en el cual se obtuvo
11% de disminucin, lo cual podra haber sido causado por
efecto de competencia entre poblaciones nativas presentes en
las columnas o de interaccin de los inculos BC y BSR, hay
reportes de fallas en los procesos de bioaugmentacin debido
a competencia entre poblaciones (Tompson et al. 2005, Van
Veen et al. 1997). Por otro lado, la degradacin de cianuro
observada en la muestra control podra deberse a la actividad
de poblaciones observadas presentes naturalmente en el relave
de cianuracin y que podran haberse incrementado por la
adicin del acetato de sodio o el cido fosfrico. Finalmente,
se pudieron haber generado factores fsico qumicos adversos
para el inoculo BC por las condiciones de trabajo en columnas.
En las pruebas realizadas con los relaves 2 y 3, se comprob
induccin de las BSR en el relave 3, por la adicin de lactato de
sodio 1 mM en un tiempo ms corto que cuando se emplearon
concentraciones menores, la cual sera la dosis adecuada para
ese relave; lo que demostrara que dependiendo del relave, la
adicin de nutrientes para incrementar la actividad microbiana
y la biorremediacin de relaves de cianuracin es una opcin
fcil de aplicar, econmica y de establecimiento sencillo.
Cada relave de cianuracin tiene caractersticas fsico-qumi-
cas especfcas, lo cual condicionara la presencia de diferentes
poblaciones naturales microbianas en cada relave que habra que
tener en consideracin para la aplicacin de inculos. Asimis-
mo, habra que considerar la realizacin de un estudio a largo
plazo ya que ha sido demostrado por Zagurya et al. (2004) que
los relaves de cianuracin son ambientes cambiantes debido a
cambios naturales de pH y poblacin, por lo que habra que
comprobar el efecto de las adiciones de nutrientes y/o inculos
en las diferentes condiciones producidas.
Aunque tenemos un mejor entendimiento sobre el papel de
la disponibilidad de nutrientes para acelerar los procesos micro-
bianos, an debemos de caracterizar las poblaciones microbianas
envueltas en el proceso de biorremediacin de estos relaves y
determinar el papel de otros nutrientes en el control del proceso
de biorremediacin estudiado.
Agradecimientos
El presente trabajo fue realizado gracias al apoyo del Concytec
(Proyecto PROCYT 236-2008).
Prueba
Inicio de Produccin de H
2
S
en el relave 3 (Das)
Control Negativo
Lactato de
Sodio
0,5 mM 16
1 mM 8
2 mM 8
Acetato de
Sodio
0,5 mM Negativo
1 mM Negativo
2 mM Negativo
Tabla 5. Efecto de la adicin de Lactato o acetato en la induccin
de bacterias sulfato reductoras en el relave 3. La tabla muestra el
resultado de pruebas por triplicado.
192
Hurtado & Berastain
Rev. peru. biol. 19(2): 187 - 192 (Agosto 2012)
Literatura citada
American Public Health Association (APHA), American Water Wor-
ks Association (AWWA) y Water Environment Federation
(WEF). 1999. Standard Methods for the Examination of
Water and Wastewater 20
th
Edition. pp 4-176, 4-50.
Atlas R. & R. Bartha. 2002. Ecologia Microbiana y Microbiologia
Ambiental. 2da.edn. Prentice Hall Hispanoamericana.
Pp. 562-597.
Bechard G., S. Rajan & W.D.Gould. 1993. Characterization of a
Microbiological Process. In: BioHydrometallurgical te-
chnologies.. A.E. Torma, M.L. Apel & C.L. Brierley, eds.
The Minerals, Metals & Material Society.
De Bolder P. 2003. Metal Bioavailability and speciation in a wetland
tailings Repository Amended with Biosolids Compost,
Wood Ash and sulfate. J. Environ. Qual. 32: 851-864
Freyre Antuane. 2000. Biodegradacin de desechos de cianuracin
producidos en la industria minera. Tesis para optar el t-
tulo de Licenciado en Ciencias con mencin en Qumica.
Universidad Peruana Cayetano Heredia.
Gerhardt P., R.G.E. Murray, R,N, Costilow et al.1981. Manual of
Methods for General Bacteriology. American Society for
Microbiology. Pp 456-458.
Knowles 1976. Microorganisms and cyanide. Bacteriol. Rev.40:652-
680.
Lawrence R & B. Marchant. 2006. Developments and new appli-
cations for biogenic sulphide reagent in hydrometallurgy
and Mineral processing. Hidroprocess, Santiago. Chile.
Luque-Almagro V. 2005. Bacterial degradation of cyanide and its
metal Complexes under alkaline conditions. Appl. Environ.
Microbiol. 71:940-947.
Muyzer, G. & A.J.M. Stams. 2008. The ecology and biotechnology
f sulphate-reducing bacteria. Nature Rev. Microbiol.
6:441-454
Rivera J. & J. Hurtado. 2001. Solid media for isolation of ferrocyani-
de degrading microorganisms. Proceedings of International
Biohydrometallurgical Symposium IBS-2001, Brasil.
Scheeren P.J.H, R.O. Koch, N. Buisman et al. 1992. New Biological
Tretament for heavy metal contaminated ground water.
Trans. Instn. Min. Metall. ( Sect. C. Mineral Process. Extr.
Metall.) 101:C190-C199
Scherr K.E, M. M. Nahold, W. Landschbauer & A.P.Loibner. 2011.
Sequential application of electron donors and humic acids
for the anaerobic bioremediation of chlorinated aliphatic
hydrocarbons, New Biotech. 29: 116-125
Sitte J., D.M. Akob,C. Kaufman et al. 2010. Microbial Links between
Sulfate Reduction and Metal Retention in Uranium and
Heavy Metal-Contamined Soil. Appl. Eviron. Microbial.
76:3143-3152
Starkey RL. 1938. A study of spore formation and other morpho-
logical characteristics of Vibrio desulfuricans. Arch.
Mikrobiol. 9: 268-304,
Strattan D.J. 2010 .Quantifying the expression of dissimilatory
sulfte reductase as a metric for sulfate reduction rates.
Thesis Submitted in partial fulfllment of the requirements
for the degree of Master of Science in Geology in the
Graduate College of the University of Illinois at Urbana-
Champaign.<http://www.ideals.illinois.edu/bitstream/
handle/2142/16814/3_Strattan_Derik.pdf?sequence=5>
Acceso 23/12/2011.
Thompson I.P., C.J.Van der Gast, L.N. Ciric & A.C. Singer 2005.
Bioaugmentation for bioremediation: the challenge of
strain selection. Env. Microbiol.7:909-915.
Van Veen J.A., L.S. van Overbeek & J.D. van Elsas. 1997. Fate and
activity of microorganisms introduced into soil. Microbial
Molecul. Biol. Rev. 61:121-135
Velarde C. 1992. La minera en el Peru. Editores Tecnicos Aso-
ciados. Peru
White C. & G.M. Gadd. 1996. Mixed sulphate-reducig bacterial
cultures for bioprecipitation of toxic metals: factorial and
response-surface analysis of the effects of dilution rate,
sulphate and substrate concentration. Microbiology. 142:
2197-2205.
Witlock James. 1990. Biological process in mining: Present and
future. Western regional Conference. Lead S.D. Sept.
Zagureya G.J., K. Oudjehanib & L. Deschenesb. 2004. Characteriza-
tion and availability of cyanide in solid mine tailings from
gold extraction plants. Sci. Total Environ. 320:211-224
193
Dosis nica de ciclofosfamida disminuye la calidad espermtica y el epitelio germinal
Rev. peru. biol. 19(2): 193 - 198 (August 2012)
Rev. peru. biol. 19(2): 193 - 198 (Agosto 2012)
Facultad de Ciencias Biolgicas UNMSM
ISSN 1561-0837
Dosis nica de ciclofosfamida disminuye la calidad espermtica y
el epitelio germinal masculino en ratones
Lyonal Acosta
1, 3
*, Vctor Nez
1, 3
, Jonathan Vsquez
1,2
, Jos Pino
1
y Betty Shiga
1
Unique cyclophosphamide doses in male mice decrease the spermatic
quality and germinal epithelium
1 Laboratorio de Reproduccin y
Biologa del Desarrollo, Departa-
mento de Zoologa. Facultad de
Ciencias Biolgicas. Universidad
Nacional Mayor de San Marcos
(UNMSM). Lima, Per. Av. Vene-
zuela cdra. 34, Lima, Per. Tel.
+511 619 7000 extensin 1529,
fax: +511 4649110. PO BOX 11-
0058. Lima 11.
2 Laboratorio de Fisiologa de la
Reproduccin Animal. Instituto de
Investigacin en Ciencias Biolgi-
cas Antonio Raimondi, Facultad
de Ciencias Biolgicas, UNMSM.
Lima, Per.
3 REPROLAB PER SAC. Lima
Per.
* Autor para correspondencia:
Email Lyonal Acosta:
acostaclg@gmail.com
Presentado: 08/02/2012
Aceptado: 20/09/2012
Publicado online: 10/11/2012
Resumen
Ciclofosfamida (CP) es un agente alquilante comnmente utilizado como frmaco antineoplsico e inmuno-
supresor. El uso de CP, en el tratamiento de cncer es limitado debido a su severa toxicidad. El objetivo del
presente estudio fue determinar el efecto de una nica dosis de CP (220 mg/kg peso corporal) administrada
intraperitonealmente despus de 7 das, sobre parmetros espermticos y el epitelio germinal masculino en
ratones. Se utiliz machos maduros de la cepa C57BL divididos en 2 grupos (n= 10), Grupo Control (C): NaCl
0,9 % y Grupo Tratamiento (T): CP a 220 mg/kg pc en dosis nica y despus de 8 das fueron eutanizados.
Se registr el peso corporal y de rganos reproductivos: testculos, epiddimos y conducto deferente, se eva-
luaron la concentracin, movilidad y morfologa espermtica e histologa testicular. Se encontraron diferencias
signifcativas (Prueba t- student p<0,05) en el peso corporal (T: 20,181,9 g vs C: 23,351,2 g); peso testicular
(T: 59,715,40 mg vs C: 72,902,8 mg), cola del epiddimo (T: 9,831,54 mg vs C: 14,302,20mg, conducto
deferente (T: 8,270,80 mg vs C: 10,610,76 mg respectivamente), concentracin (T: 9,21,9 vs C: 14,43,3
x 10
6
espermatozoides/mL), Movilidad progresiva (T: 25,986,4% vs C: 42,965,2%). En la morfologa esper-
mtica no se observ diferencias signifcativas (Prueba t- student p<0,05), sin embargo en anomalas de la
cola y espermatozoides inmaduros se observ un aumento signifcativo (T vs. C: 36,247,8% vs 31.739,1%;
27,442,1% vs 18,518,6%; respectivamente) y a su vez se observ severos daos en los tbulos seminferos.
Concluimos que una dosis nica de CP (220mg/kg pc) disminuye la calidad reproductiva de ratones macho.
Palabras clave: Ciclofosfamida; ratn C57BL; toxicidad; fertilidad; anlisis seminal.
Abstract
Cyclophosphamide (CP) is an alkylating agent commonly used as antineoplastic and immunosuppressive
drug. The use of CP in the treatment of cancer is limited due to its severe toxicity. The aim of this study was to
determine theeffectof a single dose of CP (220 mg/kg b.w.) intraperitoneally administered after 7 days,on sperm
parameter and male germinal epithelium in mice. We used C57BL mature male mice divided into 2 groups (n=
10); Control Group (C): 0.9% and NaCl Treatment group (T): CP to 220 mg/kg cw in single dose. After 8 days
all the mice were euthanized. The weight of the body and reproductive organs: testis, epididymis and vas defe-
rens, were registered. Concentration, motility and sperm morphology and testicular histology were evaluated.
Signifcant differences were found in body weight (student t-test p<0.05) (T: 20.181.9 g vs C: 23.351.2 g;
testicular weight (T: 59.715.40 mg vs C: 72.902.8 mg), epididymis tail weight (T: 9.83 1.54 vs C: 14.30
2.20, vas deferens weight (T: 8.270.80 mg vs C: 10.610.76 mg respectively), concentration (T: 9.21.9 vs. C:
14.43.3 x 10
6
sperm / mL), progressive motility (T: 25.986,4% vs C: 42.965.2%). There wasnt signifcant
differences in sperm morphology (student t-test p<0.05), but there was a signifcant increase in abnormal tail
and immature sperm (T vs. C: 36,247,8% vs 31.739,1%; 27,442,1% vs 18,518,6%; respectively) and also
severe damage seminiferous tubules was observed. We conclude that single dose of CP (220mg/kg b.w.)
decreased the reproductive quality of male mice.
Keywords: Cyclophosphamide; C57BL mouse; toxicity; fertility; semen analysis.
Introduccin
Ciclofosfamida (CP) [N, N-bis (2-cloroetil) tetrahidro-2H-1,
3,2 oxaza - fosforin-2-amino 2-oxido] es un agente alquilante
comnmente utilizado como frmaco antineoplsico e inmu-
nosupresor. La citotoxicidad de la CP es mediada por la alqui-
lacin del ADN en la posicin N7 de guanina y la formacin
de cross-links del tipo ADN-ADN y ADN-protena, as como
quiebres en una de las hebras del ADN (Hemminki & Kallama
1986, Crook et al. 1986).
La ciclofosfamida afecta clulas en divisin; por ejemplo,
genera apoptosis celular en la lnea espermtica (Crook et al.
1986, Chowdhury 1987, Brinkworth & Nieschlag 2000) e
interfere con las actividades mitticas y meiticas de la hilera
germinal. Su mecanismo de accin consiste en alquilar el ADN
alterando la estructura de los cidos nucleicos, por lo tanto la
divisin celular no se realiza adecuadamente (Elangovan et al.
2006). En el testculo, CP interrumpe la meiosis antes de la
etapa de paquiteno, produciendo dao genotxico (Barton et
al. 2003). Cuando los alquilantes se utilizan en bajas dosis, la
recuperacin de la espermatognesis podra ocurrir dentro de los
3 aos post-tratamiento; sin embargo, a altas dosis la infertilidad
podra prolongarse o incluso ser permanente.
Pacientes varones con cncer tratados con CP presentan una
mayor incidencia de oligozoospermia y azoospermia (Charak et
al. 1990, Kenney et al. 2001), adems se ha demostrado per-
turbaciones en la secrecin de gonadotropinas, dao testicular
y bajos niveles de testosterona en la sangre (Hoorweg-Nijman
et al. 1992).
194
Acosta et al.
Rev. peru. biol. 19(2): 193 - 198 (Agosto 2012)
La inyeccin nica de CP durante la formacin de esperma-
tocitos de ratn resulta en la aparicin de translocaciones hereda-
bles (Sotomayor & Cumming 1975) e incrementa la incidencia
de microncleos (Tates 1992, Lahdetie 1983, 1988). El estudio
morfomtrico en las diferentes fases del ciclo espermatognico
indica que la toxicidad en el testculo puede ser detectada a
partir de los 7 das despus de una administracin nica de
CP (100 mg/kg pc) en ratas (Matsui et al. 1995). Estudios en
ratas y ratones han confrmado que CP en dosis nica semanal
durante 5 semanas disminuye el peso testicular, la concentracin
espermtica y produce oligozoospermia transitoria (Elangovan
et al. 2006).
El estudio de los efectos de CP es muy importante debido a
su amplio uso en quimioterapias y a sus efectos secundarios no
deseados asociados con periodos variables de infertilidad (Trasler
et al. 1988). El objetivo del presente estudio fue determinar el
efecto despus de 7 das de slo una dosis de CP (220mg/kg pc)
administrada intraperitonealmente sobre la calidad espermtica
y el epitelio germinal masculino en ratones.
Materiales y mtodos
Animales.- Se emplearon ratones (Mus musculus) de la cepa
C57BL, machos (N= 20) de 8 a 10 semanas de edad con peso
promedio de 21,71,9 g, provenientes del Bioterio del Labo-
ratorio de Reproduccin y Biologa del Desarrollo, Facultad
de Ciencias Biolgicas de la Universidad Nacional Mayor de
San Marcos. Los animales fueron mantenidos bajo condiciones
estndar de bioterio (24 27 C de temperatura, fotoperodo
de 14 h luz: 10 h oscuridad) y alimentados con dieta balanceada
para animales (Purina Per) y agua ad libitum.
Diseo experimental.- Los ratones fueron divididos en dos
grupos: el grupo Tratamiento (T) (n= 10), recibi intraperi-
tonealmente (ip) una dosis de Ciclofosfamida (NEOPHOS
- Lima) de 220 mg/kg [dosis equivalente a la teraputica en hu-
manos (750 mg/kg)] (Elangovan et al. 2006) y el grupo Control
(C) (n= 10), que recibi ip una dosis de solucin salina 0,9%.
Al octavo da despus del tratamiento se registr el peso
corporal y los animales fueron eutanizados por dislocacin cer-
vical. Se pesaron: testculos, epiddimos y conductos deferentes.
Se obtuvieron espermatozoides de la cola del epiddimo para
registrar la movilidad, concentracin y morfologa espermtica.
Los testculos fueron separados para histologa.
Obtencin de epiddimos y espermatozoides.- Cada uno
de los epiddimos obtenidos fueron lavados con tampn fosfato
salino (PBS pH 7,4) a 37C, se realizaron varios cortes en la cola
del epiddimo y luego los espermatozoides se liberaron, perma-
neciendo durante 10 minutos en 0,5 ml de cultivo Flushing
(MediCult, Copenhagen, Dinamarca), el contenido esperm-
tico se recuper en su totalidad en tubos de polipropileno de
1,5 mL (Axygen Scientifc).
Evaluacin de la movilidad espermtica.- Una muestra
espermtica fue colocada sobre una lmina portaobjetos que
se encontraba a 37 C y observada a 400X en un microscopio
ptico de contraste de fases (Carl Zeitz Jena, San Francisco,
U.S.A). Este procedimiento fue repetido 2 veces, los resultados
presentados son el promedio de ambas evaluaciones, siendo
evaluados al menos 100 espermatozoides. Se consider que un
espermatozoide presentaba Movilidad Progresiva (MP), cuando
presentaba desplazamiento, Movilidad No Progresiva (MNP) a
aquellos que no se desplazaban y slo tenan un movimiento
in situ e Inmviles (I) los que no presentaban ningn tipo de
movimiento (WHO 2010).
Medicin de concentracin espermtica.- La concentra-
cin espermtica se midi usando una dilucin 1:20, 10 L de
la muestra se diluy con 190 L de solucin fjadora (WHO
2010), posteriormente se coloc 10 L del diluido en una cmara
de Neubauer, y se contaron los espermatozoides dentro de los
campos defnidos en la cmara (5 cuadrantes), mediante el uso
de un microscopio ptico de campo claro a 400X; la cantidad
de espermatozoides observados se multiplic por el factor co-
rrespondiente (10
6
). La concentracin de espermatozoides fue
expresada en millones/L.
Evaluacin de la morfologa espermtica.- Los esperma-
tozoides fueron expandidos mediante un frotis sobre lminas
portaobjetos se dej secar a temperatura ambiente, se fjaron
con paraformaldehdo por 20 minutos y se colorearon con
Eosina-Y al 5%. Por individuo fueron visualizados y analizados
250 espermatozoides bajo un microscopio ptico de campo claro
con objetivo de inmersin (100X). Se consider como anormal,
aquel espermatozoide que presentaba alteraciones morfolgicas
en cabeza, pieza intemedia, cola o en presencia de ms de una
alteracin. Espermatozoides con gota citoplasmtica fueron
considerados inmaduros.
Histologa.- Para el estudio histolgico, ambos testculos
fueron trozados y fjados en Bouin alcohlico por 16 horas,
posteriormente fueron lavados y deshidratados en una batera de
alcoholes e incluidos en Paraplast. Se realizaron cortes seriados
de 6 m, los cuales fueron coloreados con Hematoxilina
Eosina-Y. La observacin se realiz mediante un microscopio de
campo claro (Carl Zeiss Jena). Las fotomicrografas se tomaron
con una cmara digital.
Anlisis estadstico.- Los resultados fueron evaluados usando
el paquete estadstico SPSS 17,0 para Windows. Se realiz el
test de Shapiro Wilk y Leneve para comprobar normalidad y
homocedasticidad de los datos. Las diferencias entre los grupos
se analizaron mediante la prueba t-Student. Los resultados son
expresados como media EE (error estndar), n= 10. Un valor
de p<0,05 fue considerado estadsticamente signifcativo.
Resultados
Peso corporal y rganos reproductivos.- Se encontr dife-
rencias signifcativas entre el peso corporal y los rganos repro-
ductivos evaluados (testculo, epiddimo y conducto deferente).
Los resultados se resumen en la Tabla 1.
Movilidad y concentracin espermtica.- La concentra-
cin espermtica disminuy signifcativamente en el grupo T
(p<0,05) (Tabla 2). En la movilidad se observa una diferencia
signifcativa (p<0,05) en los espermatozoides con movilidad
progresiva (MP) e inmoviles (I) del grupo T con respecto al
control (25,986,4 vs 42,965,2 y 50,876,3 vs 36,054,9, res-
pectivamente).No se observ diferencias signifcativas (p<0,05)
en los espermatozoides con movilidad no progresiva (MNP) (T:
26,681,8 vs C: 25,384,3) (Fig. 1).
Morfologa.- Las evaluaciones morfolgicas no mostraron
diferencias signifcativas (prueba t-Student, correccin arco seno,
p>0,05) en el porcentaje de espermatozoides normales entre el
grupo T y el grupo C (Tabla 2). Al evaluar la distribucin de
195
Dosis nica de ciclofosfamida disminuye la calidad espermtica y el epitelio germinal
Rev. peru. biol. 19(2): 193 - 198 (August 2012)
anormalidades (cantidad neta de ocurrencias por cada anorma-
lidad observada) se observ un aumento signifcativo (p<0,05),
dentro de la fraccin anormal, de espermatozoides inmaduros
(gota citoplasmtica) (T: 27,442,1 vs C: 18,518,6) y anorma-
lidades de la cola (T: 36,247,8 vs 31.739,1). No se observ
diferencias signifcativas (p<0,05) en los espermatozoides con
anormalidades de la cabeza (T: 23,486,2 vs C: 23,915,9)
y anormalidades de la pieza intermedia (T: 46,196,8 vs C:
50,596,5) (Fig. 2)
Anlisis Histolgico.- Se observ una severa alteracin
morfolgica producida en los tbulos seminferos de ratones
tratados con ciclofosfamida 220 mg/kg por peso corporal,
entre estas alteraciones se puede mencionar una reduccin en
el dimetro del tbulo seminfero (Fig. 3A), prdida parcial del
tejido intersticial y en el nmero de clulas espermatogoniales
(Fig. 3C) con respecto a los controles (Figs. 3B, 3D).
Discusin
El presente estudio evalu si una sola dosis de CP (220 mg/
kg pc) produce algn efecto negativo sobre la fsiologa repro-
Control (Na Cl 0,9%) Ciclofosfamida (220 mg/kg pc)
Ratones examinados 10 10
Peso corporal inicial (g) 21,400,7 21,901,0
Peso corporal fnal (g) 23,351,2 20,181,9*
Peso del testculo (mg) 72,902,8 59,71 5.40*
Peso cabeza + cuerpo del epiddimo (mg) 19,250.9 16,960.7*
Peso de la cola del epiddimo (mg) 14,302,20 9,831,54*
Peso del conducto deferente (mg) 10,610,76 8,270,80*
Los valores son expresados como media SD.
*P<0.05 en comparacin con el grupo control.
Tabla 1. Efecto de la CP sobre el peso corporal, peso de testculo, epiddimo y conducto deferente entre el grupo Control (C) y Tratamiento
(T). pc: peso corporal.
Figura 1. Efecto de la CP a una sola dosis sobre la movilidad esper-
mtica. MP: Movilidad progresiva; MNP: Movilidad no progresiva; I:
Inmovilidad. pc: peso corporal. Los valores son expresados como
media SD. *P<0,05 en comparacin con el grupo control.
Control
(Na Cl 0,9%)
Ciclofosfamida
(220 mg/kg pc)
N de ratones examinados 10 10
Concentracin
espermtica (x 10
6
/mL)
14,403,3 9,21,9*
Morfologa Normal (%) 39,721,3 38,124,5
Morfologa Anormal (%) 60,281,3 61,884,5
Los valores son expresados como media SD.
*P<0.05 en comparacin con el grupo control.
Tabla 2. Efecto de la CP a una sola dosis sobre la concentracin y
Morfologa espermtica. pc: peso corporal
Control NaCl 0,9% Ciclofosfamida 220 mg/kg pc
0
10
20
30
40
50
60
Cabeza Pieza
intermedia
Cola Gota
citoplasmtica
(%)
Anormalidad morfolgica
*
*
0
10
20
30
40
50
60
MP MNP I
(%)
Movilidad espermtica
Control NaCl 0,9% Ciclofosfamida 220 mg/kg pc
*
*
Figura 2. Efecto de la CP a una sola dosis sobre los diferentes
tipos de anormalidades morfolgicas del espermatozoide. pc: peso
corporal. Los valores son expresados como media SD. *P<0.05 en
comparacin con el grupo control.
196
Acosta et al.
Rev. peru. biol. 19(2): 193 - 198 (Agosto 2012)
ductiva, espermiacin y permanencia del espermatozoide en el
epiddimo. Trasler et al. (1986) y Brinkwort y Nieschlag (2000)
utilizaron dosis de CP (5,1 mg/kg pc) por 9 y 11 semanas
respectivamente y Da et al. (2002) proporcion 5,0 mg/kg pc
por 28 das, sin observar modifcaciones signifcativas respecto a
los pesos corporales y los pesos de rganos reproductivos como
testculos y epiddimos; sin embargo, Elangovan et al. (2006)
inyect una dosis ms alta de CP (200 mg/kg pc) y observ
despus de 7 das una disminucin del peso corporal y pesos
de los testculos y epiddimos, concordando con los resultados
reportados en este trabajo inyectando una sola dosis de CP
(220 mg/kg pc) y evaluado al octavo da (Tabla 1, p<0,05).
La disminucin de los pesos corporales con una dosis alta
(CP 200 mg/kg) nos sugiere existira un efecto sistmico en
el individuo y que dosis menores durante tiempos prolongados
(Trasler et al. 1986, Brinkwort & Nieschlag 2000) seran ms
adecuadas en los tratamientos antineoplsicos con CP. Los
testculos, epiddimos y prstata son rganos dependientes de
andrgenos (Trentacose et al. 2001). Cuando administramos
una dosis nica de CP (220 mg/kg pc) observamos una dismi-
nucin signifcativa en los pesos de los testculos y epiddimos
por lo que concluimos que una inyeccin de CP a alta dosis
estara ejerciendo actividad antiandrognica a nivel gonadal
o a nivel del eje hipotlamo-hipfsis, resultados observados
tambin con dosis menores continuas (Poblador et al. 1990,
Elangovan 2006, Resvanfar et al. 2008). El peso de los testcu-
los depende de la masa de las clulas madres espermatognicas
(Katoh et al. 2002), por lo que CP (220 mg/kg pc) estara
ejerciendo dao sobre las mismas (Fig. 3).
La espermatognesis es un proceso altamente sincronizado
y dinmico que se lleva a cabo en los tbulos seminferos, en
los ratones este ciclo tiene una duracin de 35 das (Oakberg
1956). El tiempo de evaluacin post CP en este estudio (7
das) indica su accin sobre la espermiacin y el transporte
en el epiddimo (Pia-Guzmn et al. 2004). La inyeccin
nica de CP ejerce efectos anti reproductivos, disminuyendo
los valores de movilidad y concentracin espermtica. La
disminucin de la MP y el aumento de los espermatozoides
inmviles observados (Fig. 1) nos permite sugerir que CP
podra estar impidiendo la secrecin de factores proteicos en
el epiddimo cuya funcin es completar la maduracin de los
espermatozoides (Bedford 1979).
Figura 3. Tbulos seminferos de ratn BalbC57 tratados con CP (220mg/kg pc) a una sola dosis: A (100X) y C (630X); Controles: B (100X)
y D (630X). Note la disminucin del tamao de los tbulos (fecha).
197
Dosis nica de ciclofosfamida disminuye la calidad espermtica y el epitelio germinal
Rev. peru. biol. 19(2): 193 - 198 (August 2012)
No se observ aumento en la cantidad neta de espermato-
zoides anormales (Tabla 2, p<0,05) en consecuencia del breve
periodo de evaluacin de los espermatozoides post- inyeccin de
CP (7 das), de esta manera se explica porqu la administracin
de CP con dosis de 6 mg/kg pc por 28 das (Resvanfar et al.
2008) y 50 mg/kg pc por 35 dias (Arencibia et al. 2009, Valiente
et al. 2010) si incrementan el porcentaje de espermatozoides con
formas anormales. Sin embargo, observamos mayor cantidad de
espermatozoides con presencia de gota citoplasmatica, indicio
de inmadurez del espermatozoide (Fig. 2), fenmeno observado
en fuerte concordancia con el tiempo de evaluacin que abarca
la etapa de espermiacin. El estres oxidativo inducido por CP
se traduce en el aumento de celulasapoptoticas (Elangovan et
al. 2006), este factor y el dao al ADN durante el transito en
el epiddimo pueden ser los responsables del incremento en el
porcentaje de espermatozoides inmaduros.
La concentracin espermtica presenta una relacin directa
con la disminucin del peso testicular (Katoch et al. 2002) y la
apoptosis de las espermatogonias por accin de la CP disminuye
el nmero de espermatozoides (Brinkwort & Nieschlag 2000)
siendo esto confrmado en nuestro anlisis histolgico (Fig. 3C).
En nuestro estudio observamos una disminucin signifcativa en
la concentracin espermtica (Tabla 2, p<0,05), concordando con
los resultados de Valiente et al. (2010) y Elangovan et al. (2006)
quienes suministraron CP 50 mg/kg pc por 35 das y CP 200 mg/
kg pc una vez por semana durante 5 semanas, respectivamente.
Al realizar la histologa de los testculos se observ una reduc-
cinen el dimetro de los tbulos seminferos y en el nmerode
estratos de clulas germinales de los ratones tratados con CP,
siendo estos resultados similares a los obtenidos por Elangovanet
al. (2006),quienes demuestran que una dosis de CP a 200 mg/kg
pc en ratones una vez por semana durante cinco semanas presenta
el mismo dao teniendo efectos sobre la espermatognesis y la
fertilidad. Estas observaciones se deberan a los bajos niveles de
gonadotropinas y testosterona en el plasma sanguneo despus
de la inyeccin de CP (Means 1975, Chowdhury 1979, Elan-
govan et al. 2006) concordando con el efecto antiandrognico
que postulamos anteriormente para CP.
Basados en los resultados del presente estudio concluimos que
una dosis nica de CP (220 mg/kg pc) altera la espermiacin y
permanencia en el epiddimo disminuyendo la concentracin
y movilidad espermtica y a su vez daa el epitelio seminfero a
travs de la disminucin del dimetro de los tbulos seminfe-
ros y el nmero de estratos de clulas germinales de los ratones
tratados con CP. Hay que destacar que este es el primer estudio
que indica que una sola dosis de CP en alta concentracin causa
dao a nivel de la calidad reproductiva y en consecuencia sobre
la fertilidad en ratones.
Literatura citada
Arencibia D.F., L.A. Rosario & D. Curveco. 2009. Frecuencia espon-
tnea e inducida de anomalas en la morfologa de la cabeza
del espermatozoide en ratones NMRI. Retel 20(1):2-14.
Barton T.S., A.J. Wyrobek, F.S. Hill, et al. 2003. Numerical Chro-
mosomal Abnormalities in Rat Epididymal Spermatozoa
Following Chronic Cyclophosphamide Exposure.Biol.
Reprod. 69(4): 1150-7
Bedford J.M. 1979. Evolution of the sperm maturation and sperm
storage functions of the epididymis. Baltimore: Urban and
Schwarzenberg. pp. 721.
Brinkworth M. & E. Nieschlag. 2000. Association of cyclophospha-
mide-induced male-mediated, fetal abnormalities with
reduced paternal germ-cell apoptosis. Mutat. Res. 447
(2): 149-154.
Charak B., R. Gupta, P. Mandrekar, et al. 1990. Testicular dysfunction
after cyclophosphamide-vincristine-procarbazine-pred-
nisolone chemotherapy for advanced Hodgkins disease.
A long-term follow-up study. Cancer 65(9): 19031906.
Chowdhury A. 1987. Effect of pharmacological agents on male
reproduction.AdvContracept. Deliv. Syst. 3(4): 347-352.
Crook T.R., R.L. Souhami& A.E. Mclean. 1986. Cytotoxicity, DNA
cross-linking, and single strand breaks induced by activa-
ted cyclophosphamide and acrolein in human leukemia
cells. Cancer Res. 46(10): 50295034.
Das U., M. Mallick, J. Debnath, et al. 2002.Protective effect of
ascorbic acid on cyclophosphamide-induced testicular
gametogenic and androgenic disorders in male rats.Asian
J. Androl. 4(3): 201207.
Elangovan N., T.J. Chiou, W.F. Tzeng, et al. 2006. Cyclophosphami-
de treatment causes impairment of sperm and its fertilizing
ability in mice. Toxicology 222(1-2): 60 - 70.
Ghosh D., U.B. Das, S. Ghosh, et al. 2002. Testicular gametogenic
and steroidogenic activities in cyclophosphamide treated
rat: a correlative study with testicular oxidative stress.
Drug Chem. Tox. 25(3): 281292.
Hemminki K. & S. Kallama S. 1986. Reactions of nitrogen mustards
with DNA. IARC Sci. Publ. 78: 55-70.
Hoorweg Nijman J.J., H.A. Delemarre-van de Waal, F.C. de Waal,
et al. 1992. Cyclophosphamide-induced disturbance of
gonadotropin secretion manifesting testicular damage.
Acta Endocrinol. 126(2): 143148.
Katoh C., S. Kitajima, Y. Saga, et al. 2002. Assessment of quantitative
dual-parameter fow cytometric analysis for the evalua-
tion of testicular toxicity using cyclophosphamide and
ethinylestradiol treated rats. J. Toxicol. Sci. 27(2): 8796.
Kenney L., M. Laufer, F. Grant, et al. 2001. High risk of infertility
and long term gonadal damage in males treated with high
dose cyclophosphamide for sarcoma during childhood.
Cancer 91(3): 613621.
Lahdetie J. 1983. Micronuclei induced during meiosis by ethyl
methanesulfonate, cyclophosphamide and dimethylben-
zanthracene in male rats. Mutat. Res. 120(4): 257260.
Lahdetie J. 1988. Induction and survival of micronuclei in rat sperma-
tids.Comparison of two meiotic micronucleus techniques
using cyclophosphamide.Mutat. Res. 203(1): 4753.
Manda K. & A.L. Bhatia. 2003. Prophylactic action of melatonin
against cyclophosphamide-induced oxidative stress in
mice. Cell Biol. Toxicol. 19(6): 367372.
Matsui H., K. Mitsumori, K. Yasuhara, et al. 1995. Morphological
evaluation of cyclophosphamide testicular toxicity in rats
using quantitative morphometry of spermatogenic cycle
stages. J. Toxicol. Sci. 20(4): 407-414.
Means A.R. 1975. Biochemical effects of follicle stimulating hormo-
nes on the testis. In: R.O. Greep and E.B. Ashwood, eds.
Handbook of Physiology, Section 7, Endocrinology. Ame-
rican Physiological Society, Washington, USA. : 203219.
Oakberg E.F. 1956. Duration of spermatogenesis in the mouse and
timing of stages of the cycle of the seminiferous epithe-
lium. Amer. J. Anat. 99(3): 507-516.
Pia-Guzmn B., M.J. Sols-Heredia, B. Quintanilla-Vega. 2005.
Diazinon alters sperm chromatin structure in mice by
phosphorylating nuclear protamines. Toxicol. Appl. Phar-
macol. 202(2): 189-98.
Poblador M.S., C. Rojas, A. Raya, et al. 1990. The effects of cyclo-
phosphamide on the gonadotrophic cells of the normal rat.
Histol.Histopathol. 5(2): 173-80.
198
Acosta et al.
Rev. peru. biol. 19(2): 193 - 198 (Agosto 2012)
Rezvanfar M., R. Sadrkhanlou, A. Ahmadi, et al. 2008. Protection of
cyclophosphamide-induced toxicity in reproductive tract
histology, sperm characteristics, and DNA damage by an
herbal source; evidence for role of free-radical toxic stress.
Hum. Exp. Toxicol. 27: 901-910.
Sotomayor R. & R. Cumming. 1975. Induction of translocations by
cyclophosphamide in different germ-cell stages of male
mice cytological characterization and transmission. Mutat.
Res. 27(3): 375-388.
Tates A. 1992. Validation studies with the micronucleus test for early
spermatids of rats. A tool for detecting clastogenicity of
chemicals in differentiating spermatogonia and spermato-
cytes.Mutagenesis. 7(6): 411-419.
Trasler J.M., B.F. Hales, B. Robaire. 1986. Chronic low dose cy-
clophosphamide treatment of adult male rats: effect on
fertility, pregnancy outcome and progeny. Biol. Reprod.
34(2): 275283.
Trasler JM., L. Hermo, B. Robaire. 1988. Morphological changes
in the testis and epididymis of rats treated with cyclo-
phosphamide: a quantitative approach. Biol. Reprod.
38(2): 463-479.
Trentacoste S.V., A.S. Friedmann, R.T Youker, et al. 2001. Atrazine
effects on testosterone levels and androgen-dependent
reproductive organs in peripubertal male rats. J. Androl.
22(1):142-8.
Valiente A.M., Y.R. Salgado, N.S Prez, et al. 2010. Efecto de la
ciclofosfamida en la morfologa del espermatozoide del
raton.Retel 34: 1-24.
World Health Organization (WHO). 2010. Standard procedures. In:
T.G. Cooper, eds, Laboratory manual for the Examination
and processing of human semen, 5th edn. Switzerland.
199
Chemical composition of Yuca de monte, apoDanthera biflora
Rev. peru. biol. 19(2): 199 - 203 (August 2012)
Rev. peru. biol. 19(2): 199 - 203 (Agosto 2012)
Facultad de Ciencias Biolgicas UNMSM
ISSN 1561-0837
Chemical composition of Apodanthera bifora, a Cucurbit of the dry
forest in northwestern Peru
Daniel Clark
1
*, Maribel Tupa
2
, Andrea Bazn
2
, Lily Chang
3
, Wilfredo L. Gonzles
2
Composicin qumica de Apodanthera bifora, una cucurbitcea del bosque
seco del noroeste peruano
1 Unidad de Genmica, Facultad
de Ciencias y Filosofa, Universidad
Peruana Cayetano Heredia.
2 Laboratorio de Ecologa Evoluti-
va, Facultad de Ciencias y Filosofa,
Universidad Peruana Cayetano
Heredia.
3 Departamento de Ciencias Exac-
tas, Facultad de Ciencias y Filoso-
fa, Universidad Peruana Cayetano
Heredia.
*Autor para correspondencia.
Facultad de Ciencias y Filosofa,
Universidad Peruana Cayetano
Heredia. Avenida Honorio Delgado
430, San Martn de Porres, (Apar-
tado Postal 4314, Lima 100), Per.
Email Daniel Clark:
daniel.clark@upch.pe
Email Maribel Tupa:
trdu7@yahoo.es
Email Andrea Bazn:
maryandrea17@hotmail.com
Email Lily Chang:
lily.chang@upch.pe
Email Wilfredo Gonzles:
wilfredo.gonzales@upch.pe
Presentado: 17/02/2012
Aceptado: 11/08/2012
Publicado online: 10/11/2012
Abstract
The dry forest of northwestern Peru is the habitat of numerous native species that have received little atten-
tion beyond taxonomic purposes. Here we report the chemical composition of the roots and seeds of a set
of accessions of Apondanthera bifora, a wild Cucurbit with potentialities as a food resource. The roots show
features that are comparable to those from cassava roots and potato tubers: a high content of carbohydrates
on a dry matter basis (77.5 84.9%), with starch representing around 20% of the total dry matter. The seeds
fat content on a dry matter basis (22.22 39.37%) falls within the percentages found in oily seeds such as
saffower, whereas the protein content (21.37 29.06%) is similar to that of fax and sunfower. The fatty acid
profle of the seeds oil shows a predominance of polyunsaturated acids, in particular linoleic acid (43.81%),
making it comparable to that of cotton oil. Our results confrm the nutritional value of Apodanthera bifora and
set the ground for its use in domestication and food security programs.
Keywords: Yuca de monte; food security; dry forest; starch; oily seed; comestible root.
Resumen
El bosque seco del noroeste del Per es hbitat de numerosas especies nativas que, fuera de su taxonoma,
han sido poco estudiadas. En este trabajo se reporta la composicin qumica de las races y semillas de un
grupo de accesiones de Apodanthera bifora, una cucurbitcea silvestre con potencial nutricional. Las races
muestran una composicin comparable a la de la yuca y la papa: un alto contenido de carbohidratos en base
al peso seco (77,5 - 84,9%), entre los cuales destaca el almidn con alrededor de 20% del peso seco. El con-
tenido de grasas de la semilla en base al peso seco (22,22 - 39,37%) est dentro del rango descrito en semillas
oleaginosas como las de crtamo, mientras que el contenido de protenas (21,37 - 29,06%) es similar al hallado
en semillas de lino y girasol. En el perfl de cidos grasos del aceite de la semilla se observa un predominio
de los cidos grasos poliinsaturados, en particular del cido linoleico (43,81%), que lo hace comparable al del
aceite de algodn. Los resultados confrman el valor nutricional de Apodanthera bifora y sienta las bases para
su domesticacin y aprovechamiento en programas de seguridad alimentaria.
Palabras clave: Yuca de monte; seguridad alimentaria; bosque seco; almidn; semilla oleaginosa; raz co-
mestible.
Introduction
Neotropical seasonal dry forests are ecosystems characterized
by precipitations of less than 1600 mm/year, with at least 5 6
months of less than 100 mm rainfall (Pennington et al. 2000). In
southwestern Ecuador and northwestern Peru, an area with such
characteristics spans 86859 km
2
of the coastal belt and shows a
high degree of endemism (Aguirre et al. 2006). While taxonomic
classifcation and geographic distribution of plant species within
this area have been addressed by several investigators over the
years (Svenson 1946; Ferreyra 1983; Aguirre et al. 2006), their
phenology, reproductive strategies, genetic diversity, and chemi-
cal potentialities for application as food resources or in industrial
processes have received little attention, in spite of the uses given
to many of those species by the local population (Bussmann &
Sharon 2006; Bussmann & Sharon 2009; Wilfredo Gonzles,
unpublished observations).
Apodanthera bifora Cogn. (locally known as yuca de monte)
is a perennial herbaceous vine belonging to the Cucurbitaceae
family that is native to the Peruvian dry forest (Macbride 1937;
Brack 1999; Jrgensen & Len 1999; Mostacero et al. 2002).
Te aerial parts of the plant (vegetative and reproductive) grow
from December through March, during the raining season.
Upon arrival of the dry season, they dry out leaving a tuber-
ous root with latent buds buried in the soil, from which new
aerial parts will grow in the following raining season (Wilfredo
Gonzles, unpublished observations).
Like other wild species of the Cucurbitaceae family such as
Cucurbita foestidissima (DeVeaux & Shultz 1985), Cucumeropsis
mannii, Cucurbita maxima, Cucurbita moschata, Lagenaria sicer-
aria, Cucumis sativus (Achu et al. 2005), or Melothria pendula
(ArazateFernndez & GrennCascales 2002), A. bifora has
some features that make it a potential resource for food security
and/or food industry. Its roots accumulate starch, are comes-
tible and often used by the local population to feed livestock.
Occasionally, though, they are also consumed by people who
incorporate them as part of their meals. In addition, its fruit, a
modifed berry or pepo, contains approximately 20 seeds with
signifcant amounts of extractable oil (Wilfredo Gonzles, un-
published observations). Nevertheless, no studies addressing the
chemical composition of the roots and seeds of A. bifora have
been carried out to date.
An efort to make a full appraisal of the potentialities of A.
bifora is being carried out in our laboratory, which includes
studies on the phenology, reproductive biology, genetic diversity,
200
Clark et al.
Rev. peru. biol. 19(2): 199 - 203 (Agosto 2012)
and chemical composition of roots and seeds, taking advantage
of a set of accessions from northwestern Peru. In the present
article we show the relevant results on the chemical and nutri-
tional value of A. bifora roots and seeds, emphasizing on the
variability observed. Te results are discussed with regard to the
values reported for domesticated species of known nutritional
and commercial importance.
Material and methods
Samples. Plant material was collected from 15 locations
in the departments of Tumbes, Piura, and Lambayeque, in
northwestern Peru (Table 1). Te locations were at least 5 km
away from each other. Approximately 10 kg/location of tuberous
roots from wild adult plants were collected in Tumbes (locations
1 and 2), Piura (locations 4 11), and Lambayeque (locations
12 14) between May and June 2009, at the end of the wet
season, when the local population harvest them. Seeds were
obtained from fruits of wild adult plants in Tumbes (location
2), Piura (locations 3 9), and Lambayeque (location 15) in
March 2009, when the fruits were mature. Both roots and seeds
were stored in paper bags at room temperature until use.
Processing of root samples. For each location, 2 kg of
undamaged roots were washed thoroughly with running water
and their cortex removed so as to leave the inner edible parts.
Te latter were washed with distilled water, chopped into small
pieces, and ground in a food processor. Te homogeneous paste
was divided into portions of approximately 40 g that were ac-
curately weighed and stored at -20 C until analysis.
Processing of seed samples. Clean seeds were frst weighed
and then desiccated in an oven at 103 C for 17 h. After weigh-
ing the desiccated seeds, the dry matter content was calculated
and the seeds ground in a mortar to determine the fat and
protein content.
Determination of chemical composition of roots.
Dry matter content: Te dry matter content of the root paste
was determined according to the AOAC Method 22.018 (AOAC
1980). Determinations were made in 6 replicates of 5 g each, 3 of
which were utilized later to quantify the lipid content and the 3
remaining to determine the ash content. Results were expressed
as percentage (w/w) of the samples fresh weight.
Ash content: Te ash content was determined in triplicate
using dehydrated samples (see previous section) according to
AOAC Method 7.009 (AOAC 1980). Between 2 5 g of dehy-
drated material were used per replicate and the results expressed
as percentage (w/w) of the samples dry weight.
Crude protein content: Te crude protein content was mea-
sured in triplicate according to AOAC Method 47.021 (AOAC
1980), with 2 g of root paste per replicate. A standard conver-
sion factor (6.25) was used to express the protein content as a
percentage (w/w) of the samples dry weight.
Lipid content: Te amount of lipids contained in the roots
was determined in triplicate with previously dehydrated samples
(see section 2.4.1.) according to AOAC Method 155 7.056
(AOAC 1980). Results were expressed as percentage (w/w) of
the samples dry weight.
Crude fber content: Te crude fber content was quantifed
in triplicate according to AOAC Method 7.061 (AOAC 1980).
Two grams of defatted sample were used per replicate and the
results were expressed as percentage (w/w) of the samples dry
weight.
Available carbohydrate content: Te percentage of available
carbohydrate in roots was estimated by subtracting from 100
the sum of water, ash, protein, lipid, and crude fber.
Ca and P contents: Samples from ashing were digested for
30 min with 6 mL of concentrated hydrochloric acid under
heating. Te digest volume was brought to 50 mL with double
distilled water in a volumetric fask and used for Calcium and
Phosphorus determinations in triplicate.
Calcium was quantifed in 3 replicates by the permanganate
titration method described by James (1999), with slight modi-
Location Department Coordenate S Coordenate W Altitude Root analysis Seed analysis
1 Tumbes 3 32' 57.0012" 80 18' 27" 0 x
2 Tumbes 3 38' 21.9984" 80 24' 14.0004" 10 x x
3 Piura 5 06' 12.999" 80 34' 13.0008" 119 x
4 Piura 5 08' 41.9994" 80 32' 48.0006" 176 x x
5 Piura 5 12' 47.9988" 80 23' 40.9992" 188 x x
6 Piura 5 13' 21" 80 35' 57.9984" 154 x x
7 Piura 5 22' 58.9974" 80 16' 50.9988" 243 x x
8 Piura 5 32' 39.0006" 80 28' 18.0006" 246 x x
9 Piura 5 34' 51.999" 80 14' 34.0008" 238 x x
10 Piura 5 44' 28.9998 80 18' 38.9988" 199 x
11 Piura 5 46' 22.998" 80 12' 15.9978" 209 x
12 Lambayeque 6 17' 17.9982" 79 52' 58.9974" 68 x
13 Lambayeque 6 22' 55.9986" 80 1' 5.9982" 115 x
14 Lambayeque 6 15' 35.9994" 80 4' 19.9986" 139 x
15 Lambayeque 5 47' 21.9978" 79 59' 39.9984" 166 x
Table 1. Collection sites of the specimens included in this study.
201
Chemical composition of Yuca de monte, apoDanthera biflora
Rev. peru. biol. 19(2): 199 - 203 (August 2012)
fcations (AOAC Method 7.096, 1980). Briefy, the diluted ash
digest is brought to pH 4.5 and an excess of ammonium oxalate
is added to precipitate calcium as calcium oxalate. After several
washing rounds with 2% ammonium hydroxide, oxalic acid is
liberated from the precipitate by adding sulfuric acid and titra-
tion of the acid is carried out afterwards with 0.01 M potassium
permanganate at 70 C to a persistent pink color. Results were
expressed as milligrams of calcium per 100 g of dried sample.
Phosphorus was determined in 3 replicates with a HI 83225
photometer (Hanna Instruments) and phosphorus reagents
HI 93706A0, HI 93706B0, and HI 9370601 (HANNA
Instruments) according to the manufacturers instructions. Te
kit is based on the amino acid method, adapted from Standard
Method for the Examination of Water and Wastewater (1999).
Te reaction between phosphorus and the reagents gives a blue
color in the sample. Results were expressed as milligrams of
phosphorus per 100 g of dried sample.
Determination of protein, lipid content, and fatty acid
composition of seeds. Total protein and fat content in seeds
were quantifed in triplicate according to AOAC Methods
47.021 and 7.056, respectively (AOAC 1980) and expressed
as percentage of dry matter. Te fatty acid composition of the
oil obtained was determined by gas chromatography in a HP
6890 apparatus (Agilent Technologies, Palo Alto, CA). Briefy, a
capillary column Omegawax 100 (15 m, 0.10 mm i.d., 0.10 m
flm thickness) was used for the separation of fatty acid methyl
esters. Te initial temperature was 140 C for 5 min, raised to
280 C at a rate of 40 C/min, and kept at 280 C for 2 min.
Te split ratio was 1:200 and the carrier gas hydrogen kept at
a constant fow rate of 50 cm/s. Te injector temperature was
250 C and the detector temperature 280 C. One L of sample
was injected and the running time was 60 min.
Statistical analysis. To assess whether the chemical compo-
sition of the tuberous root is afected by the site of collection, a
nested analysis of variance (ANOVA) was performed considering
as factors the department (Lambayeque, Piura or Tumbes) and
the location (nested in the department). Descriptive statistics
were used to address the seeds chemical features of interest such
as the fat and protein content, and the fatty acid composition of
the oil obtained. Te mean, range (minimum and maximum),
and coefcient of variation (standard deviation/mean) are re-
ported for each seed trait.
Results and discussion
Chemical composition of the root. Te results of the
proximate analysis carried out with roots from 13 locations
in northwestern Peru are given in Table 2. Te content of cal-
cium and phosphorus was also included. With the exception
of the fber content, all the parameters show some degree of
variation at the department level (Table 3). It is worth noting
the high percentage of fat and calcium content found in roots
from Lambayeque as compared with those from Tumbes and
Piura. Te percentage of fat in Lambayeque is 2.3 2.5 fold
higher than those in Tumbes and Piura, whereas the amount
of calcium shows a 1.4 fold increase with respect to Tumbes
(Table 2). Diferences were also evident within departments at
the location level, with fber content as the sole exception again
(Table 3). Te reason for this apparent uniformity in the fber
content among accessions is unclear at present and a search in
the scientifc literature did not reveal a similar trend in other
comestible storage roots such as cassava (Charles et al. 2005).
Notable variations were observed among all the populations in
the fat percentages (from 0.1 to 9.2%), as well as in the amounts
of calcium (from 37.3 to 132.7%) and phosphorus (from 69 to
182.3%). All these variations were concentrated in Piura, with
Dry mass % Ash % Fiber % Fat % Protein % Calcium mg% Phosphorous mg%
Departments
Piura 26.2 0.3 a** 4.9 0.1 a 0.7 0.1 NS 3.1 0.5 a 6.4 0.3 a 95.6 6.3 a 112.8 8.1 a
Lambayeque 23.0 0.6 b 5.6 0.2 b 1.1 0.1 NS 7.8 0.9 b 8.0 0.7 b 107.2 4.6 b 91.3 4.4 b
Tumbes 23.7 1.2 b 4.3 0.3 c 1.8 1.0 NS 3.4 0.5 a 7.9 1.6 ab 67.7 8.8 c 110.9 4.0 ab
Locations
1 19.8 0.3 e 4.5 0.1 ab 2.9 1.9 NS 3.5 0.1 abd 4.5 1.1 a 49.3 2.7 b 117.8 5.7 bc
2 27.6 0.3 bc 4.1 0.5 a 0.7 0.1 NS 3.3 1.1 ab 11.3 0.2 b 86.0 6.4 ad 104.0 0.4 bc
3 27.0 0.4 b 4.4 0.1 ab 0.4 0.0 NS 1.1 1.1 ab 8.8 0.1 ab 115.1 8.5 acd 80.8 4.0 bc
4 28.9 0.4 c 4.1 0.1 a 0.2 0.2 NS 2.0 1.1 ab 5.6 0.1 a 132.7 2.4 c 182.3 3.7 ad
5 27.0 0.2 b 4.8 0.1 ab 0.4 0.1 NS 1.1 1.0 ab 5.5 0.1 a 88.4 4.9 ad 69.0 0.6 c
6 27.6 0.4 bc 4.8 0.1 ab 0.6 0.0 NS 0.1 0.1 ae 4.7 0.2 a 37.3 5.9 b 112.4 7.4 bc
7 24.5 0.2 af 4.8 0.1 ab 0.9 0.1 NS 6.4 0.6 dbe 6.4 0.4 ab 98.0 13.0 ad 100.4 2.9 bc
8 25.0 0.1 a 5.3 0.2 abc 2.0 0.2 NS 3.4 0.6 abd 6.9 0.9 ab 110.0 14.0 ac 136.0 40.0 bd
9 27.0 0.3 b 4.4 0.1 ab 0.6 0.2 NS 4.4 0.5 abc 6.4 0.3 ab 70.8 3.4 bd 119.0 11.0 bc
10 22.4 0.2 df 6.2 0.1 c 0.7 0.1 NS 5.6 1.5 abc 6.7 2.8 ab 113.2 4.4 acd 102.0 2.4 bc
11 20.5 0.5 e 5.4 0.3 b 1.0 0.2 NS 8.3 0.4 cd 8.5 2.5 ab 124.7 2.5 acd 78.2 8.4 bc
12 23.0 0.5 f 6.0 0.4 c 0.9 0.0 NS 9.2 1.9 ce 7.5 0.4 ab 97.8 1.7 acd 100.9 3.2 bc
13 25.6 0.2 a 5.4 0.1 b 1.3 0.4 NS 5.9 1.5 abc 7.8 0.2 ab 99.2 4.2 acd 94.9 3.2 bc
*Values represent the average of 3 replicates the standard deviation, except for Dry mass, whose average was calculated from 6 replicates.
**Different letters indicate signifcant differences (P<0.05) among values in a nested ANOVA.
Table 2. Chemical composition (on a dry matter basis) of Apodanthera bifora roots from 13 locations in northwestern Peru*
202
Clark et al.
Rev. peru. biol. 19(2): 199 - 203 (Agosto 2012)
the exception of the fat percentage. Still, its variation in Piura
was considerable (from 0.1% to 6.4%, Table 2). While part of
this variability might be explained by phenotypic plasticity under
diferent environmental conditions (precipitation, temperature,
soil composition), genetic diferences cannot be ruled out as an
ISSRbased estimation of the genetic diversity in the sampled
area resulted in a signifcant degree of population structure
(unpublished results). Te lower variability found in Tumbes
and Lambayeque might have been determined by the smaller
number of locations sampled.
Interestingly, the carbohydrate content of A. bifora roots
is high (77.5 84.9%) and comparable to those reported for
cassava roots (94%; USDA, National Nutrient Database for
Standard Reference) or potato tubers (86.94%; Jimnez 2005).
Tis attribute is considered an important nutritional feature
of comestible storage roots/tubers, and our results confrm the
nutritional value of the A. bifora roots as a source of carbo-
hydrates. Preliminary results with a few accessions indicated
that starch represents approximately 20% of the dry matter, a
percentage that is lower to that obtained with cassava (around
30%; Luna et al. 2006), but higher than the percentage re-
ported for potato (13 15%; Lisinska & Leszczynski 1989).
We are currently confrming these results by assessing further
accessions, as well as characterizing the starch by physicochemi-
cal methods.
A somewhat unexpected fnding was that, although low (3.1
7.8%), the fat content is 4 11 fold higher than of cassava
(USDA, National Nutrient Database for Standard Reference)
and 2 5 fold that of potato (Jimnez 2005). As for the rest of
attributes, the dry weight (23 26.2%) and the protein content
(6.4 8%) are comparable to those found in potato (26.8%
and 7.46%, respectively; Jimnez 2005), but clearly diferent
from those in cassava (40.3% and 3.4%; USDA, National
Nutrient Database for Standard Reference); ashes content were
comparable to those of cassava or potato (Charles et al. 2005;
Jimnez 2005); fber (0.7 1.8%) was lower than values reported
for cassava and potato (4.5% and 2.6%, respectively; USDA,
National Nutrient Database for Standard Reference; Jimnez
2005); phosphorous levels (91.3 112.8 mg%) were between
those found in cassava (66.9 mg%; USDA, National Nutrient
Database for Standard Reference) and potato (194 mg%; Jim-
nez 2005), while calcium levels (67.7 107.2 mg%) were higher
than those of cassava and potato (39.7 mg% and 22.4 mg%,
respectively; USDA, National Nutrient Database for Standard
Reference; Jimnez 2005).
Taken together, our results are consistent with the use of
A. bifora root as a source of carbohydrates in the regular diet
and its inclusion as part of food security programs, especially
in northwestern Peru. Further testing of the starch is currently
under way to assessing its potential for industrial applications.
Protein, lipid content, and fatty acid composition of
seeds. Te protein and fat content of seeds from 9 locations
sampled are shown in Table 4. Te fat content in Piura was
notably higher than that in Tumbes and Lambayeque, while the
amount of protein tended to be higher in Tumbes with respect to
the other 2 departments (data not shown). However, the fact that
only 1 accession from Tumbes and Lambayeque was evaluated
does not allow drawing defnitive conclusions. Te range of fat
content in all accessions (22.22 39.37%) falls within the values
reported for other oily seeds, in between peanut (36 56%) and
cotton (15 22%), and very similar to safower (25 40%).
Te protein percentage (21.37 29.06%) is somewhat lower
Dry mass (%) Ash (%) Fiber (%) Oil (%) Protein (%) Carbohidrates (%) Calcium mg% Phosphorous mg%
DF F
Department 2 117.33*** 21.66*** 3.02NS 27.18*** 3.90* 31.87*** 20.77*** 3.51*
Location
(Department)
10 66.71*** 6.42*** 1.54NS 3.43* 3.72* 3.98* 16.05*** 6.13***
a
F values are reported
***P < 0.0001, **P < 0.001, *P < 0.05, NS P > 0.05; Degrees of freedom: Error = 26; Total = 36
Table 3. Nested analysis of variance (ANOVA) of the chemical variables taking as factors the department (Tumbes, Piura or Lambayeque) and
the locations (nested in the departments)
a
Range
Mean (%) Min. Max. SD
a
CV
b
Protein
c
23.99 21.37 29.06 2.58 0.11
Fat
d
31.79 22.22 39.37 5.78 0.18
Fatty acids
C14:0 0.08 0.00 0.14 0.05 0.63
C16:0 17.41 15.40 20.57 1.74 0.10
C16:1 0.10 0.00 0.18 0.07 0.65
SUM C16 17.52 15.56 20.57 1.70 0.10
C18:0 14.11 13.14 14.88 0.68 0.05
C18:1 22.29 18.00 25.19 1.96 0.09
C18:2 43.81 39.38 46.56 2.30 0.05
C18:3 0.00 0.00 0.00 0.00 0.00
SUM C18 80.21 78.23 83.51 1.88 0.02
C20:0 0.73 0.61 0.88 0.09 0.12
C20:1 0.00 0.00 0.00 0.00 0.00
C22:0 0.19 0.00 0.99 0.32 1.72
SUM C20 0.92 0.61 1.71 0.36 0.39
Others 1.22 0.00 2.16 0.87 0.72
Saturated 32.52 29.53 34.90 2.13 0.07
Mono unsaturated 22.40 18.00 25.36 2.01 0.09
Poli unsaturated 43.81 39.38 46.56 2.30 0.05
a
Standard deviation
b
Coeffcient of variation
c
Sampled size = 7
d
Sampled size = 9
Table 4. Fat and protein content (on a dry matter basis), and fatty acid
composition of Apodanthera bifora seeds.
203
Chemical composition of Yuca de monte, apoDanthera biflora
Rev. peru. biol. 19(2): 199 - 203 (August 2012)
than that reported for soja (35 44%), and comparable to fax
(20 24%) and sunfower (18 25%) (Lpez Bellido 2002).
Te fatty acid composition as determined by gas chromatog-
raphy is shown in Table 4. Tere was a slight predominance of
polyunsaturated over saturated fatty acids, whereas the mono-
unsaturated were clearly the less abundant. When examining
the individual fatty acid content, the percentage of linoleic acid
(43.81%) stands out, followed by oleic (22.29%), palmitic acid
(17.41%), and stearic acid (14.11%). Te percentage of other
fatty acids was below 1%, and therefore considered not signif-
cant. Te overall composition of the oil is comparable to that
of cotton (http://www.scientifcpsychic.com/ftness/fattyacids1.
html), with a strong dominance of n6 over n3 fatty acids. As
a matter of fact, no linolenic acid could be detected in the oil
of any of the accessions analyzed.
While the fat and fatty acid composition are compatible with
the use of the A. bifora seedderived oil for human consump-
tion, the short life of the mature fruit in wild specimens makes
difcult to build up sufcient starting material for an industrial
approach.
Concluding remarks. Because of its chemical composi-
tion, the root of A. bifora emerges as a useful resource for food
security in arid areas such as the dry forest of northwestern Peru;
in addition, its starch content opens the possibility of industrial
applications whose viability is currently being assessed. Te
seedderived oil also shows interesting characteristics for human
consumption as a food additive, although the short life of the
mature fruit curtails its production at an industrial scale. Domes-
tication eforts might prove useful to overcome this limitation.
In that regard, our investigations have revealed a phenotypic and
genetic diversity in the population studied that is promising for
the future development of a domestication program.
Acknowledgements
We thank Anbal Caldern, Jenny Rojas, David de Las Casas,
Luis Espinoza, Adn Yarlequ, Lucinda Prado, Yolmer Gutir-
rez, Adan Zea, Alberto Valdera, Jos Cedillo, Dante Alemn,
the Comunidad Campesina Jos Ignacio Tvara Pasapera
(Chulucanas, Morropn), the Empresa Comunal de Servicios
Agropecuarios Cucungara S.R.L. (Catacaos, Piura), and the
Asociacin para la conservacin de la Biodiversidad y Desarrollo
Sustentable de los Bosque Secos del Morante (ACBIODESA,
Catacaos, Piura) for their assistance in the feld labors. We
also thank the Laboratory of Quality Control at Alicorp S.A.
for their valuable assistance in the analysis of fatty acids, the
Peruvian Ministry of Agriculture for authorizing sample collec-
tion (Permit N 00582009AGDGFFSDGEFFS), and the
herbarium of the Universidad Peruana Cayetano Heredia where
the specimens collected are preserved. Tis work was supported
by the Programa de Ciencia y Tecnologa (FINCyT), Contrato
N002-FINCyT-PIBAP-2009.
Literature cited
Achu M.B., E. Fokou, C. Tchigang, M. Fotso & F.M. Tchouanguep.
2005. Nutritive value of some Cucurbitaceae oilseeds
from different regions in Cameroon. African Journal of
Biotechnology. (4): 13291334.
Aguirre Z., R. LinaresPalomino & L.P. Kvist. 2006. Woody spe-
cies and vegetation formations in seasonally dry forests of
Ecuador and Peru. Arnaldoa. (13): 324346.
AOAC. 1980. Offcial methods of Analysis of AOAC, 13AOAC
(1980). Offcial methods of Analysis of AOAC, 13th
edition. Washington D.C, USA. Association of Offcial
Analytical Chemists.
ArazateFernndez A.M. & G.N. GrennCascales. 2002. Contri-
bucin al conocimiento del pepinillo silvestre (Melothria
pendula L.). Ciencia Ergo Sum. (9): 7886.
Brack A. 1999. Diccionario Enciclopdico de plantas tiles del Per.
Cusco, Per. Editorial Bartolom de las Casas.
Bussmann R.W. & D. Sharon. 2006. Traditional medicinal plant use
in Northern Peru: tracking two thousand years of healing
culture. Journal of Ethnobiology and Ethnomedicine. (2):
4764.
Bussmann R.W. & D. Sharon. 2009. Shadows of the colonial pastdi-
verging plant use in Northern Peru and Southern Ecuador.
Journal of Ethnobiology and Ethnomedicine. (5): 420.
Charles A.L., K. Sriroth, & T. Huang. 2005. Proximal composition,
mineral contents, hydrogen cyanide and phytic acid of 5
cassava genotypes. Food Chemistry. (92): 615620.
Clescerl L.S., A.E. Greenberg, & A.D. Eaton, eds. 1999. Standard
Method for the Examination of Water and Wastewater,
20
th
edition.
DeVeaux J.S. & E.B. Shultz. 1985. Development of Buffalo Gourd
(Cucurbita foetidissima) as a semiaridland starch and oil
crop. Economic Botany. (39): 454472.
Ferreyra R. 1983. Los tipos de vegetacin de la costa peruana. Anales
Jardn Botnico de Madrid. (40): 241256.
James C.S. 1999. Analytical chemistry of foods. Frederick, MD,
USA. Aspen Publishers, Inc.
Jimnez F.S. 2005. Caractersticas nutricionales de la arracacha (Ar-
racacia xanthorrhiza) y sus perspectivas en la alimentacin.
Red Peruana de Alimentacin y Nutricin (rPAN).
Jrgensen P.M. & S. Len, eds. 1999. Catalogue of the vascular
plants of Ecuador. Monographs in Systematic Botany from
the Missouri Botanical Garden. 75. St. Louis, MO, USA:
Missouri Botanical Garden.
Lisinska G. & W. Leszczynski. 1989. Potato Science and Technol-
ogy. London, New York. Elsevier Science Publishers Ltd.
LpezBellido L. 2002. Cultivos Industriales. Barcelona, Espaa.
MundiPrensa.
Luna W.A. & J.A. Mera. 2006. Produccin de dextrinas de yuca a
partir de almidn nativo en la rallandera Todoyuca ubicada
en el corregimiento Pescador (municipio de Caldono,
Cauca). BEng Thesis, Universidad del Cauca, Colombia.
Macbride J.F. 1937. Cucurbitaceae Flora of Peru. Field Museum
of Natural History, Botanical Series. (13)6/2: 321383.
Mostacero L.J., F.C. Meja & O.T. Gamarra. 2002. Taxonoma de
las fanergamas tiles del Per. Concytec. (1): 538539.
Pennington R.T., D.E. Prado, & C.A. Pendry. 2000. Neotropical
seasonally dry forests and Quaternary vegetation changes.
Journal of Biogeography. (27): 261273.
Svenson H.K. 1946. Vegetation of the coast of Ecuador and Peru
and its relation to that of the Galapagos Islands. II Catalog
of plants. American Journal of Botany. (33): 427498.
USDA, National Nutrient Database for Standard Reference, Release
25 (online). Basic Report, Nutrient data for 11134, Cassa-
va, raw. < http://ndb.nal.usda.gov/ndb/foods/show/2892>
[acceso: 29/10/2012]
204
Clark et al.
Rev. peru. biol. 19(2): 199 - 203 (Agosto 2012)
205
Prediciendo la distribucin de polylepiS
Rev. peru. biol. 19(2): 205 - 212 (August 2012)
Rev. peru. biol. 19(2): 205 - 212 (Agosto 2012)
Facultad de Ciencias Biolgicas UNMSM
ISSN 1561-0837
Prediciendo la distribucin de Polylepis: bosques Andinos vulnerables
y cada vez ms importantes
Brian R. Zutta
1,2
, Phillip W. Rundel
1
, Sassan Saatchi
2
, Jorge D. Casana
3
, Paul Gau-
thier
4
, Aldo Soto
5
, Yessenia Velazco
6
, y Wolfgang Buermann
7
Predicting Polylepis distribution: vulnerable and increasingly important
Andean woodlands
1 Center for Embedded Networked
Sensing, University of California
Los Angeles, 3563 Boelter Hall, Los
Angeles, CA 90095-1596, EEUU.
Email Brian R. Zutta:
bzutta@ucla.edu
Email Phillip W. Rundel:
rundel@lifesci.ucla.edu
2 Jet Propulsion Laboratory, Cali-
fornia Institute of Technology, 4800
Oak Grove Drive, Pasadena, CA
91109, EEUU.
saatchi@congo.jpl.nasa.gov
3 Parque Nacional Huascarn,
Instituto Nacional de Recursos
Naturales, Federico Salyrosas N
555, Huaraz, Per.
4 Research School of Biology.
The Australian National University,
Canberra, ACT 0200. Australia
paul.gauthier@anu.edu.au
5 Centro de Datos para la Con-
servacin, Universidad Nacional
Agraria La Molina, Av. La Molina
S/N, Apartado 12-056, Per.
aldo_soto@lamolina.edu.pe
6 Department of Biological Scien-
ces, California State University Los
Angeles, 5151 State University Dri-
ve, Los Angeles, CA 90032, EEUU.
yvelazc@calstatela.edu
7 Center for Tropical Research,
Institute of the Environment, Uni-
versity of California Los Angeles,
La Kretz Hall, Suite 300, Box
951496, Los Angeles, CA 90095-
1496, EEUU.
buermann@ucla.edu
Presentado: 23/05/2012
Aceptado: 23/08/2012
Publicado online: 13/11/2012
Resumen
Los bosques de Polylepis son recursos vitales para la conservacin de la biodiversidad y funciones hidrolgicas,
la cual se ver alterada por el cambio climtico a nivel mundial desafando la sostenibilidad de las comunidades
locales. Sin embargo, estos ecosistemas andinos de gran altitud son cada vez ms vulnerables debido a la
presin antropognica como la fragmentacin, deforestacin y el incremento en el ganado. La importancia para
predecir la distribucin de bosques nativos ha aumentado para contrarrestar los efectos negativos del cambio
climtico a travs de la conservacin y la reforestacin. El objetivo de este estudio fue desarrollar y analizar
los modelos de distribucin de dos especies, Polylepis sericea y P. besseri, que forman bosques extensos a
lo largo de los Andes. Este estudio utiliz el programa Maxent, el clima y capas ambientales de una resolucin
de 1 km. El modelo de distribucin previsto para P. sericea indica que la especie podra estar situada en una
variedad de hbitats a lo largo de la Cordillera de los Andes, mientras que P. besseri se limitaba a las grandes
alturas del sur de Per y Bolivia. Para ambas especies, los metros de elevacin y la temperatura son los factores
ms importantes para la distribucin prevista. El perfeccionamiento del modelo de Polylepis y otras especies
andinas utilizando datos de satlites cada vez ms disponibles al pblico demuestran el potencial para ayudar
a defnir las reas de diversidad y mejorar las estrategias de conservacin en los Andes.
Palabras claves: distribucin, Maxent, MODIS, Polylepis, sensores remotos, QSCAT, WorldClim.
Abstract
Polylepis woodlands are a vital resource for preserving biodiversity and hydrological functions, which will be
altered by climate change and challenge the sustainability of local human communities. However, these high-
altitude Andean ecosystems are becoming increasingly vulnerable due to anthropogenic pressure including
fragmentation, deforestation and the increase in livestock. Predicting the distribution of native woodlands has
become increasingly important to counteract the negative effects of climate change through reforestation and
conservation. The objective of this study was to develop and analyze the distribution models of two species that
form extensive woodlands along the Andes, namely Polylepis sericea and P. weberbaueri. This study utilized
the program Maxent, climate and remotely sensed environmental layers at 1 km resolution. The predicted
distribution model for P. sericea indicated that the species could be located in a variety of habitats along the
Andean Cordillera, while P. weberbaueri was restricted to the high elevations of southern Peru and Bolivia. For
both species, elevation and temperature metrics were the most signifcant factors for predicted distribution.
Further model refnement of Polylepis and other Andean species using increasingly available satellite data
demonstrate the potential to help defne areas of diversity and improve conservation strategies for the Andes.
Keywords: Maxent, MODIS, Polylepis, QSCAT, remote sensing, species distribution modeling, WorldClim.
Introduccin
Cada vez ms vulnerables, los bosques de Polylepis (Rosaceae)
son un ecosistema Andino importante para la proteccin de la
biodiversidad y la mitigacin de los efectos del cambio climtico
local. El gnero Polylepis incluye aproximadamente 27 especies
(Mendoza & Cano 2011, Kessler & Schmidt-Lebuhn 2006)
que forman bosques de hoja perenne con poblaciones muy
fragmentadas a lo largo de las alturas de los Andes tropicales
y subtropicales. Se estima que quedan menos del 10% de su
extensin original en las regiones altas de Bolivia y Per (Fjelds
& Kessler 1996) y catorce especies estn listados como vulnera-
bles (IUCN 2011). Sin embargo, estos bosques contienen una
variedad de especies endmicas y en peligro de extincin (Servat
et al. 2002) y cumplen funciones importantes hidrolgicas en
intercepcin de neblina (Fig. 1; Fjelds 2002).
Todas las especies de Polylepis son arbustos o rboles, algunos
hasta 10 m de altura y comparten caractersticas morfolgicas,
que incluyen troncos rojos y torcidos, corteza delgada y exfo-
liante, y pequeas hojas imparipinnadas. Varias especies son
Figura 1. Mezcla de bosques de Polylepis en el Parque Nacional
Huascarn, Cordillera Blanca del Per, que soporta una variedad de
epftas y es importante para la intercepcin de neblina. Foto por Autor.
206
Zutta et al.
Rev. peru. biol. 19(2): 205 - 212 (Agosto 2012)
Especie Distribucin Notas sobre ecologa
P. australis Central y nor-oeste de Argentina
(e)
Alto bosque nublado, restringida a laderas y barrancos.
P. besseri Sur Per a Bolivia De 3000-4100 m en algunas reas a menudo se forman bosques puros.
P. canoi Ayacucho, Cusco y Junn, Per
(e)
De 3350-3400.
P. crista-galli Sur de Bolivia a norte de Argentina
(v)
Por debajo de 3000 a 3900 m, favorece laderas secas en las elevaciones
ms bajas.
P. favipila Andina occidental del centro del Per
(e)
Pendientes ridas de 3600-4100 m.
P. hieronymi Sur de Bolivia hasta el norte de Argentina
(v)
Especie pionera que se encuentra por debajo 3000-3400 m y laderas
orientales.
P. incana Ecuador hasta el sur de Per
(v)
Laderas occidentales de Ecuador desde 3500-4100 m. Hasta 4200 m en
Per.
P. incarum Sur Peruano a Bolivia Principalmente en 3800-4200 m en zonas con lluvias estacionales.
P. lanata Sur Peruano a Bolivia Pendientes envueltas en neblina de 2900-4100 m.
P. lanuginosa Ecuador
(e,v)
Limitados a ambientes ridos de montaa.
P. microphylla Ecuador y aislado en Per
(v)
Aislado en el Volcn Chimborazo, Ecuador y trasplantes precolombinos
en Per.
P. multijuga Norte del Peru
(e,v)
Bosques montaosos bajos.
P. neglecta Central y sur de Bolivia
(e,v)
Encontrado en 2400-3500 m y tiende a crecer en arroyos o en lomas
expuestas.
P. pacensis Bolivia occidental
(e)
Valles inter-andinos y cordilleras occidentales desde 3200-4100 m.
P. pauta Ecuador y aislado en Per y Bolivia
(v)
Bosques de montaa hasta 4000 m, en zonas con neblina frecuentes.
P. pepei Peru central hasta el noreste de Bolivia
(v)
En la zona de transicin de los bosques montaosos hmedos a pajo-
nales de puna.
P. quadrijuga Cordilleras orientales de Colombia
(e)
Por encima de la lnea de bosque continuo hasta el pramo.
P. racemosa Per a Bolivia, y aislado en Ecuador
(v)
Lmite superior de bosques montaosos hmedos y zonas de niebla
frecuentes.
P. reticulata Ecuador y aislado en el norte y centro del Per
(v)
Se encuentran en los bosques nublados altos y algunos valles inter-
andinos.
P. rugulosa Suroeste de Per a extremo norte de Chile
(v)
Entre 3000-4600 m por can y laderas orientados sudoeste.
P. sericea** Venezuela a Bolivia
Bosques hmedos de montaa, de 2600 m hasta 4100 m en muchas
zonas del Per.
P. subsericans Centro y sur de Per
(e,v)
Encontrado en 2900-5100 m, a menudo en las partes altas de caones.
P. subtusalbida Sur de Per y Bolivia central
(e)
En las regiones con una precipitacin anual de 600 mm y una altura de
3000-4500 m.
P. tarapacana Frontera de Argentina, Bolivia, Chile, y Per
(c)
Se encuentran en alturas extremas de 4000-5200 m y paisajes ridos.
P. tomentella Bolivia a Argentina, y aislado en Per central
Ocupan una variedad de hbitats de mucha altitud con precipitaciones
de 450-800 mm.
P. triacontandra Sur de Per a Bolivia, aislado en centro de Per
Altos lmites de los bosques hmedos, a 4000 m en regiones con niebla
frecuente.
P. weberbaueri** Ecuador hasta el sur de Per
(v)
Los bosques hmedos montaosos, de 2500-4300 m.
*
De acuerdo al 2011 Lista Roja de Especies Amenazadas.
** Especies modelado.
(e)
Endmico,
(v)
Vulnerable,
(c)
Casi amenazada
Tabla 1. Descripcin de la distribucin de especies de Polylepis, notas sobre la ecologa y estado de conservacin*.
conocidas por particulares caractersticas ecolgicas dentro de
su distribucin en los Andes (Tabla 1). Por ejemplo, P. pauta se
puede encontrar en ceja de selva en Ecuador y grupos aislados
en el sur Peruano (Cierjacks et al. 2008). Polylepis tarapacana
es conocida por alcanzar altitudes hasta 5200 m en el Volcn
Sajama en Bolivia, que puede ser la elevacin ms alta para las
plantas leosas en el mundo (Garcia-Nuez et al. 2004). Polylepis
sericea es la nica especie que se encuentra en Venezuela y tiene
la distribucin ms amplia del gnero, alcanzando hasta el norte
de Bolivia (Azcar et al. 2007).
La distribucin natural de los bosques de Polylepis ha sido
debatida durante mucho tiempo como consecuencia de la actual
fragmentacin potencialmente resultante de presin de miles
de aos de poblaciones humanas en los Andes (Kessler 2002).
Durante los aos, la hiptesis alternativas han sugerido que los
bosques de Polylepis originalmente cubra la mayor parte de las
montaas andinas (Ellenberg 1979) o haban rangos limitados
por microclimas especfcos (Simpson 1986). La reconstruccin
de paleo bosques de Polylepis ha sido difcil, ya que el polen
fsil es casi idntico a la del gnero Acaena (Rosaceae) (Kessler
2002). Aunque el polen en elevaciones por encima de 4800 m
es ms probable que sea de Polylepis en lugar de Acaena que es
principalmente restringido a bosques nublados (Hanselman et al.
2005), los paleo bosques son muchas veces identifcados como
Polylepis/Acaena (Hanselman et al. 2005; Gosling et al. 2008).
Hoy en da, estos bosques enfrentan una presin de pastoreo
de ganado (Teich et al. 2005), fuego (Cierjacks et al. 2008) y
la expansin de carreteras (Purcell & Brelsford 2004), todos
los cuales han reducido la extensin de hbitat y su integridad
(Jameson & Ramsay 2007). Complicando las evaluaciones de
conservacin, se han presentado las similitudes morfolgicas de
muchas especies de Polylepis que han llevado a muchos estudios
para agruparlos como un nico tipo de bosque (Simpson 1986;
Kessler & Schmidt-Lebuhn 2006). Los Modelos de distribucin
son una tcnica que mejora continuamente para estimar el
207
Prediciendo la distribucin de polylepiS
Rev. peru. biol. 19(2): 205 - 212 (August 2012)
potencial de una especie que ocurren en zonas que no han sido
previamente estudiados (Guisan & Tuiller 2005; Hernandez et
al. 2008). Las estrategias de conservacin frecuentemente utili-
zan modelos para dirigir estudios sobre la ubicacin de especies
(Raxworthy et al. 2003; Bourg et al. 2005; Hernandez et al.
2008) e identifcar reas prioritarias para la conservacin (Wilson
et al. 2005; Hernandez et al. 2008). Estudios recientes utilizando
modelos de distribucin tocan estos temas para la conservacin
de los hbitats amenazados (Riordan & Rundel 2009) y ayudar
a vincular la distribucin gentica y geogrfca (Tomassen et
al. 2011). En general, mucho de estos modelos solo requieren
datos de localizacin y datos ambientales para realizar un anlisis
estadstico que establezca las relaciones especie-medio ambiente
y construir un modelo espacial de ocurrencia.
Cada vez ms disponibles, imgenes de satlites y relativamen-
te rpidos modelos de anlisis de distribucin pueden contribuir
con una nueva perspectiva sobre la distribucin de bosques de
Polylepis y otras especies andinas con regmenes climticos es-
trechos y distinguir las reas de prioridad para su conservacin.
El objetivo de este estudio fue modelar la distribucin potencial
de la amplia gama Polylepis sericea y el vulnerable P. weberbaueri
utilizando datos de clima y sensores remotos de alta resolucin
(1 km.). Este estudio tambin identifcar el alcance geogrfco
de reas protegidas que contienen estos bosques.
Mtodos
Especies enfocadas y localizacin.- Las dos especies de Po-
lylepis de este estudio tienen distintos rangos y ocupan diferentes
hbitats en la Cordillera Andina. Polylepis sericea (Wedd.) tiene
la ms amplia distribucin del gnero con una gama geogrfca
conocida desde Venezuela a Bolivia (Kessler 1995) muchas
veces formando bosques hmedos de montaa. Habita en una
variedad de ecosistemas de altitud desde el pramo al norte del
Ecuador hasta regiones hmedas y tibias de Bolivia, el cual tiene
temporadas ms defnidas. Polylepis weberbaueri (Pilg.) aparece
desde el centro hasta el suroeste de Ecuador, en el noreste de
Per y poblaciones aisladas ms al sur de la Cordillera Blanca y
la zona de Cuzco-Apurimac. Tambin es considerada vulnerable
debido a una reduccin en el tamao de la poblacin de 20%
en los ltimos 10 aos debido a fragmentacin y la recoleccin
de madera (IUCN 2011).
En el presente estudio, las observaciones geogrfcas (locali-
dades) representan la presencia observada de una especie. Para
mejorar la precisin, este estudio escogi las localidades para P.
sericea (N = 85, Fig. 2A) y P. weberbaueri (N = 82, Fig. 3A) del
Missouri Botanical Gardens VAST (VAScular Tropicos) base de
datos de nomenclatura (http://mobot.mobot.org/W3T/Search/
vast.htm) y Fjelds y Kessler (2004), que reuni y verifc las
localidades dentro de los pasados quince aos para evitar reas
que se puedan haber deforestado. Esto tambin asegura que las
localidades fueron seleccionadas durante la adquisicin de los
datos de sensores remotos utilizados en nuestra investigacin.
Antes de modelado, los puntos fueran superpuesta sobre las
imgenes de Google Earth (versin 4.2) para verifcar la exactitud
de cada localizacin geogrfca, basado en el las descripciones de
la fuente (por ejemplo pas, departamento, altitud, proximidad
al punto de referencia, etc.) y la visibilidad los imgenes de
moderada y alta resolucin.
Capas ambientales.- Las capas ambientales contenan ambas
capas de clima y ambientes remotamente sensoreadas producidas
en los pasados seis aos, el cual cubre Sudamrica en una reso-
lucin de 1 km (Tabla 2). Las capas del clima consisten en una
Figura 2. (A) Las locali-
dades para P. sericea (N
= 85) desde Venezuela
hasta Bolivia y (B) distri-
bucin potencial.
208
Zutta et al.
Rev. peru. biol. 19(2): 205 - 212 (Agosto 2012)
serie de variables bioclimticas, conocidas como Bioclim, obte-
nidas de la base de datos WorldClim (http://www.worldclim.
org; Hijmans et al. 2005). Los datos de Bioclim representan las
tendencias anuales de temperatura y precipitacin producidas
de un conjunto mundial de datos de estaciones de clima entre
los aos 1950-2000.
Las capas ambientales remotamente sensoreadas consisten
en mediciones pticas, microondas, y radares que miden una
gama diversa de variables ambientales. Se utiliz el producto
global de TERRA MODIS LAI (MOD15A2) de resolucin de
1 km (Myneni et al. 2002). Compuestos mensuales (32 das)
del ndice de rea de Hoja (LAI) de cuatro aos (2001-2004)
fueron formados para reducir los efectos de nubes y otros dis-
turbios atmosfricos. La cobertura porcentual del rbol (~1 km.)
del ao 2001 del MODIS vegetation continuous feld (VCF),
con una resolucin nativa de 500 m, fue tambin utilizados y
puede servir como un indicador de la distribucin espacial de
la densidad de la biomasa (Hansen et al. 2002). Se incluyeron
los datos globales del sensor de microonda QSCAT (Quick
Scatterometer) disponibles en compuestos de 3 das en 2001 en
una resolucin de 2,25 km. Las medidas son sensibles al conte-
nido de agua en la hoja, la humedad, y aspereza de la superfcie
(Kunz & Long 2005). Las medidas topogrfcas, promedio de
elevacin y la desviacin estndar, se derivaron de los datos
digitales de la elevacin de SRTM, aumentadas desde 90 m a
1 km. de resolucin.
Despus de la reduccin de datos para llegar al mnimo de
covarianza y facilitar la interpretacin de la distribucin de
las especies (ver Buermann et al. 2008), un total de 18 capas
del medio ambiente, en una resolucin de 1 km espacial, se
utilizaron inicialmente para este anlisis. El subconjunto f-
nal de 11 capas bioclimticas incluyen la temperatura media
anual, promedio de temperatura diurna, estacionalidad de la
temperatura, la temperatura mxima del mes ms caluroso, la
temperatura mnima del mes ms fro, la precipitacin anual, la
estacionalidad de precipitacin, la precipitacin de los trimestres
ms hmedos, las precipitaciones de los trimestres ms secos, la
precipitacin del trimestres ms clido, y la precipitacin de los
trimestres ms fros.El subconjunto de cinco capas de sensores
remotos incluye el mximo anual LAI, el rango anual de LAI,
el promedio anual de QSCAT y la desviacin estndar de QS-
CAT (estacionalidad), y el porcentaje de cobertura arbrea.Las
dos capas de topografa del SRTM (altura media y desviacin
estndar) tambin fueron incluidos.
Modelacin.- Nuestra investigacin requiri el uso del pro-
grama de Maxent (versin 3.2.1), el cual utiliz un algoritmo
de mxima entropa para modelar la distribucin de la especie,
encontrando la distribucin ms cercana a una distribucin
Figura 3. (A) Las localidades
para P. weberbaueri (N = 82)
desde Ecuador y poblaciones
aisladas en Per y (B) distribu-
cin potencial.
Datos Variable ecolgica Fuente Resolucin
Bioclim
Variables derivadas mensualmente de temperatura y
precipitaciones
WorldClim 1 km, aos ~1950-2000
Elevacin digital Topografa y rugosidad SRTM 30 m
Hoja de ndice de rea
Productividad neta, estacionalidad, la densidad de la
vegetacin
MODIS 1 km, cada 8 das
Porcentaje de cobertura arbrea La cubierta arbrea MODIS 500 m
Dispersmetro-retrodispersin Rugosidad, la estacionalidad, y humedad de la superfcie QSCAT 2,25 Km, cada 3 das
Tabla 2. Datos ambientales usados para el modelo de distribuciones potenciales de P. sericea y P. weberbaueri.
209
Prediciendo la distribucin de polylepiS
Rev. peru. biol. 19(2): 205 - 212 (August 2012)
uniforme a travs del rea del estudio (Philips et al. 2006).
Maxent proporciona un rango continuo de probabilidad a travs
del rea geogrfca de 0 a 1,0, dnde los valores altos indican la
oportunidad relativamente ms alta. En este estudio, los pxeles
con valores iguales o por encima de 0,5 se identifcan como reas
con una mayor probabilidad de presencia porque contienen
las condiciones bioclimticas tpicas del nicho de una especie.
El subconjunto fnal de once bioclimas y cinco capas de
sensores remotos con todas las localidades, se utilizaron para
construir los modelos de distribucin de las especies. Siguiendo
a Phillips et al.(2006), hicimos 10 modelos para cada especie,
seleccionando al azar 70% de los puntos para el modelo de capa-
citacin y 30% para las pruebas de modelo. El rea bajo la curva
ROC (Receiver Operating Characteristic) (AUC) proporcionan
una medida del rendimiento del modelo.Un valor de AUC para
un modelo puede variar de 0,5 (al azar) a 1,0 (discriminacin
perfecta). Es una pregunta importante que el uso general de
datos de herbario y la posibilidad de sesgando los resultados
puede afectar los resultados. Sin embargo, el efecto general con
Maxent es limitado (Costa et al. 2010) y usamos Google Earth
como otro nivel de verifcacin y eliminamos puntos que estaban
en alturas no razonables.
Resultados
Los valores de AUC fueron estadsticamente signifcativos
(P< 0,001, Mann-Whitney U-Test) para todas las predicciones
de distribucin de las especies de Polylepis, lo que indica que
todos los escenarios del modelo y las particiones de datos fueron
signifcativamente mejores que el azar. El valor medio de las
AUC, basado en 10 particiones al azar, para los modelos con
todas las capas del medio ambiente fue 0,996 0,003 para P.
sericea y 0,994 0,002 para P. weberbaueri.
La distribucin prevista de P. sericea se refeja con la amplia
distribucin en los Andes tropicales del oeste de Venezuela has-
ta el norte de Bolivia (Fig. 2B). El promedio de elevacin fue
3937 53 m, una precipitacin total anual de 927 32 mm y
un promedio de temperatura mnima de -2,2 0,5 C.Casi el
60% de los bosques se prev que se encuentra en el Per (Tabla
3) la mayora dentro de la Cordillera Blanca y Cordillera Hua-
yhuash y la Cordillera Occidental y Oriental de los Andes del
Per central.Ecuador contiene 20% de la extensin total, sobre
todo en las cordilleras Occidentales y Orientales.
Alrededor del 93% de P. weberbaueri fue previsto que ocu-
rriera en el Ecuador y el Per, con casi el 70% en los Andes
centrales del Per, particularmente en la Cordillera Blanca y
Cordillera Huayhuash (Fig. 3B).En el Per, la elevacin media
fue 4056 3 m, una precipitacin total anual de 745 1 mm
y un promedio de temperatura mnima de -4,4 0,02 C. Del
mismo modo, la altura media de estos bosques en el Ecuador
fue 3481 4 m, con una precipitacin total anual de 861
2 mm y un promedio de temperatura mnima de 3,4 0,03
C.Pequeas bolsas de hbitats fue provisto en regiones aisladas
de Bolivia, Colombia y Venezuela pero estos representan menos
del 4% del total de bosques potenciales.
La extensin de las reas protegidas previstas P. sericea y P.
weberbaueri son variables segn el pas.reas protegidas de Per
seran 8932 km
2
a una altura promedio de 3400 m y 6700 km
2
a
una altura promedio de 2600 m para P. sericea y P. weberbaueri,
sin embargo, esto slo representa alrededor del 12% y 9% de
los bosques protegidos, respectivamente.La reas protegidas en
los Andes Venezolanos, se prev que incluyen alrededor del 66%
de los bosques de P. sericea a una altura promedio de 5200 m y
en Bolivia incluira aproximadamente el 79% de P. weberbaue-
ri.Esto est en agudo contraste con el total de bosques previstos
para ser protegidas a lo largo de los Andes de alrededor del 19%
de P. sericea y el 13% de P. weberbaueri.
Discusin
Modelacin y actual distribucin de Polylepis.- La predic-
cin de la distribucin de la vegetacin de altura, en particular
los esenciales para la biodiversidad y la sostenibilidad, ser cada
vez ms importante para esfuerzos de conservacin en Amrica
del Sur. El manejo y restauracin de bosques de Polylepis tiene
un impacto directo en numerosas plantas endmicas y especies
asociadas (Purcell et al. 2004), y la sostenibilidad de las poblacio-
nes humanas locales dependiendo en servicios de los ecosistemas,
incluido el mantenimiento de la hidrologa y de la intercepcin
de neblina. La efcacia de los modelos de distribucin de las espe-
cies proviene de la capacidad de utilizar un nmero limitado de
observaciones con datos ambientales disponible al pblico para
predecir rpidamente las especies locales o hbitats adecuados.
Las discrepancias entre los hbitats modelados y observados,
probablemente como es el caso de Polylepis en este estudio,
muchas veces son el resultado de la intensiva presin antropo-
gnica histrica y moderna o limitada capacidad de dispersin
de los hbitats disjuntos. Se cree que el Per y Bolivia en la
actualidad contienen menos del 3% y 1%, respectivamente,
de la extensin histrica de los bosques de Polylepis (Fjelds &
Kessler 1996), que nuestros modelos de prediccin del potencial
de una extensin forestal ms grande de dos especies.La tasa de
deforestacin de bosques de Polylepis vara segn la regin. El 1%
se pierde en un plazo de 50 aos en la Cordillera de Vilcanota,
Per (Jameson & Ramsay 2007) hasta mltiples fragmentos
que se talan en menos de 12 aos en el noreste de los Andes
Bolivianos (Purcell &Brelsford 2004).Polylepis weberbaueri es
conocida por las poblaciones disjuntas en Ecuador y Per, y la
distribucin de modelado se extendi ms all de su rea de
distribucin conocida hasta pequeos bolsones en Colombia y
Venezuela.Clasifcado como especie vulnerable por la UICN
(2011), la presin antropognica sera la razn principal para
Especies
rea estimada (Km
2
)
Ven Col Ecu Per Bol Total
P. sericea 3,610 10,324 24,007 72,014 10,171 120,126
(AP) 2,395 3,103 5,020 8,932 3,498 22,948
P. weberbaueri 3,444
*
1,628
*
26,547
*
74,482 2,145 108,246
(AP) 2,262 - 3,894 6,690 1,692 14,538
* Poblaciones nativas no existen en el pas predicho.
Tabla 3. rea estimada (Km
2
) para P. sericea y P. weberbaueri y dentro de las reas protegidas (AP) para pases Andinos usando una esti-
mado conservativo.
210
Zutta et al.
Rev. peru. biol. 19(2): 205 - 212 (Agosto 2012)
la situacin corriente de poblaciones disyuntas, que incluyen
grandes desconexiones entre poblaciones de P. weberbaueri.
Otra posibilidad para muchas poblaciones disyuntas de
Polylepis son trasplantes contemporneos e histricos. Nuestros
modelos indican los bolsones de los hbitats adecuados fuera
de las reas conocidas por P. sericea y P. weberbaueri que refejan
el uso de Polylepis no nativos para la reforestacin en diferentes
regiones de los Andes.Trasplantes histricos, que han sobrevivi-
do y se reproducen con xito, puede haber causado poblaciones
atpicas (Kessler & Schmidt-Lebuhn 2006) y puede contribuir
a la frecuente ocurrencia de hibridacin entre especies a pesar
de su aislamiento (Schmidt-Lebuhn et al. 2006, 2007). En un
aspecto positivo, la facilidad de cultivo y trasplante de Polylepis
(Fig. 4) permitira una reforestacin rpida y el aumento de la
funcin hidrolgica en regiones que se espera sern ms secas
con la prdida de los glaciares.
Conservacin y restauracin de los bosques.- Tal vez los
temas de importancia en materia de conservacin en los Andes
es la ubicacin de las reas protegidas en toda la regin.reas
protegidas con reconocimiento nacional e internacional tienen
diferentes grados de alcance y efcacia, con un nmero mucho
menor que ocurren en ambientes de altura (Rodrguez & Young
2000, Soutullo et al. 2008). A raz de nuestros modelos de
prediccin para dos especies de altura, la ubicacin actual de
las reas protegidas, en Venezuela, Colombia, Ecuador, Per y
Bolivia, incluirn alrededor de 9% a casi el 80% de hbitats de
Polylepis en cada pas.Esto es indicativo de la falta de proteccin
estndar de ecosistemas de altura en toda la regin andina.El
alto endemismo y la creciente importancia de los bosques de
Polylepis para conservar la funcin del ecosistema se deben seguir
poniendo en relieve en esta regin para la conservacin en las
cuestiones del cambio climtico.
Independientemente del estado de conservacin, hay que des-
tacar que los fragmentos de bosques de todos los tamaos sirven
como hbitat esencial para la diversidad de alta elevacin. Incluso
los pequeos fragmentos con una superfcie total de bosques de
menos de 4,5 ha, dentro de la Cordillera Blanca del Per, se ha
demostrado que mantienen altos niveles de diversidad de aves
similares a los fragmentos de ms de 20 ha (Rios et al. 2011). Sin
embargo, los fragmentos pequeos deben permanecer dentro de
los 200 m de fragmentos ms grandes con el fn de mantener la
conexin funcional de especies de aves (Lloyd & Marsden 2011).
Los pastizales entre los fragmentos, un componente importante
del total de la matriz boscosa, es tambin un conductor de las
comunidades de Polylepis que utilizan mltiples hbitats (Lloyd
2008, Bellis et al. 2009) y deben ser incluidos en los esfuerzos
de conservacin que se centran en aumentar la diversidad local.
Todas estas facetas destacan la calidad de los bosques como un
factor importante en el apoyo de la mayor diversidad de especies
(Ros et al. 2011) y la retencin de la funcin ecolgica (Lloyd &
Marsden 2011). Esto tambin incluye la conservacin o el aumen-
to de la complejidad estructural de los bosques, que contienen
una mezcla de la regeneracin de los rboles maduros (Renison
et al. 2011), lo que aumentara la disponibilidad de nichos y los
recursos naturales de las diferentes etapas de la sucesin.
El estado de la diversidad gentica de las poblaciones de
bosques es un factor pasado por alto e importante a considerar
tambin. Como con la mayora de las especies de rboles tropi-
cales, la fragmentacin de los bosques de Polylepis disminuyen
signifcativamente la diversidad gentica de las poblaciones a
pesar de ser polinizada por el viento, que a menudo muestran
una menor prdida gentica de la fragmentacin (Hensen et al.
2012). Adems, las poblaciones en las elevaciones ms altas tie-
nen menor diversidad gentica que otras poblaciones (Aragundi
et al. 2011, Hensen et al. 2012) y se ha encontrado que la diver-
sidad gentica en P. multijuga era aproximadamente la mitad que
otro especies polinizadas por el viento (Quinteros-Casaverde et
al. 2012). Esto es posiblemente la resulta de los cuellos de botella
genticos y efectos fundadores durante las migraciones de ladera
del Holoceno o la corta duracin de la temporada de cultivo y
temperaturas ms fras en las elevaciones ms altas (Hensen et
al. 2012). La baja diversidad gentica en las elevaciones ms altas
tendr un impacto directo sobre la viabilidad de las primeras
poblaciones que migran con el cambio climtico. Adems, la
reforestacin con especies no nativas de Polylepis puede conducir
a la prdida local de la diversidad gentica de la hibridacin,
como otras especies relacionadas se sabe que se hibridan con
facilidad (Schmidt-Lebuhn et al. 2006). La buena noticia es que
gran cantidad de fujo de genes entre ellas y dentro de muchas
poblaciones de Polylepis existe (Schmidt-Lebuhn et al. 2006)
y la variabilidad gentica parece estar plenamente disponible
Figura 4. (A) Vivero forestal en la
regin de El Quilcayhuanca, Par-
que Nacional Huascarn, donde P.
sericea y P. weberbaueri se cultivan
para regenerar otros bosques de
la regin. (B) Parcelas de Polylepis
plntulas. Fotos de Autor.
211
Prediciendo la distribucin de polylepiS
Rev. peru. biol. 19(2): 205 - 212 (August 2012)
para la restauracin forestal en muchas reas (Julio et al. 2011).
Una posible estrategia es la recoleccin de semillas de mltiples
fragmentos grandes y de individuos separados por al menos
25 m dentro de los fragmentos para maximizar la diversidad
gentica para reforestacin (Quinteros-Casaverde et al. 2012).
Conclusiones
Hay varias implicaciones para futuras estrategias de conser-
vacin y restauracin basadas en los modelos de distribucin
de dos especies de Polylepis que son mucho ms grandes que
la distribucin actual. (1) Polylepis se puede restaurar a un rea
ms amplia. Puede existir muchas reas no disponibles para los
bosques, debido a la agricultura u otros usos, pero fragmentos
de Polylepis pequeos y continuos todava pueden promover la
biodiversidad y aumentar la funcin hidrolgica de la regin.
(2) Las especies de Polylepis se pueden plantar en muchas zonas
que son o no son nativas. El uso de poblaciones locales para la
restauracin de bosques sera preferible para evitar la hibridacin
y la prdida de diversidad gentica. (3) Nuevas reas protegidas,
privadas o pblicas, pueden ser creadas especfcamente para
Polylepis, en lugares apropiados climticamente y donde ahora
no existen. Un plan estratgico con bosques de Polylepis puede
incluir el uso sostenible de la biodiversidad de esta nuevo ecosis-
tema, como el mercado limitado de orqudeas y otras plantas, y
el ecoturismo, en particular la observacin de aves, que puede ser
conectados con un circuito de sitios arqueolgicos para promover
la conservacin de ambos (Puhakka et al. 2011).
El reciente aumento en los estudios y la atencin en los bos-
ques de Polylepis refejan la creciente importancia del manteni-
miento de este ecosistema. Sin embargo, todava quedan muchos
espacios con respecto a nuestro conocimiento de las especies y la
historia de la distribucin de los bosques, la ubicacin actual, y
la respuesta al cambio climtico. Modelizacin de la distribucin
actual y futura de todas las especies de Polylepis mejorar con la
disponibilidad de los datos ambientales de alta resolucin. Los
modelos regionales del clima, producidas especfcamente para
predecir el cambio climtico en el futuro de Amrica del Sur,
se espera que sean ms precisos que los modelos de circulacin
global que ahora se utilizan para la regin. Una alternativa
importante para la adquisicin de imgenes son las imgenes
de alta resolucin disponibles en Google Earth, que han sido
utilizados para clasifcar nuevos fragmentos de bosque en el sur-
central del Per (Benham et al. 2011) y representan un nuevo,
libre y efcaz mtodo de anlisis de bosques de gran altitud. La
susceptibilidad de estos bosques a una mayor fragmentacin y
la prdida gentica requieren una mayor proteccin y una pla-
nifcacin cuidadosa si la restauracin es sostenible para todas
las especies de Polylepis. Esperamos que la tecnologa y tcnicas
avanzadas de manera signifcativa ayuden a mejorar el futuro de
los bosques y las especies que dependen de ellos.
Agradecimientos
Agradecemos a Jorge Leal-Pinedo, Guillermo Velazco Zuta,
Kiko Edson Roncal Velasco y Liliana Zuta de Roncal por la
apreciable asistencia recibida durante los estudios de campo.
Agradecer a los revisores annimos por sus valiosos comentarios
sobre versiones anteriores del manuscrito. Apreciamos tambin
el apoyo de Juan Carlos Castro Vargas, ex-director del Parque
Nacional Huascarn, Per. Nos gustara dar las gracias asimismo,
al UCLA Latin America Center, UCLA Vavra fund, y al progra-
ma UC Pacifc Rim el cual proporcion fnanciamiento a Autor.
Literatura citada
Azcar A., F. Rada & C. Garca-Nez. 2007. Functional characte-
ristics of the arborescent genus Polylepis along a latitudinal
gradient in the high Andes. Interciencia 32: 663-668.
Bellis L.M., L. Rivera, N. Politi, E. Martn, M.L. Perasso, F. Cornell &
D. Renison. 2009. Latitudinal patterns of bird richness, di-
versity and abundance in Polylepis australis mountain forest
of Argentina. Bird Conservation International 19: 265-276.
Benham P.M., E.J. Beckman, S.G. DuBay, L.M. Flores, A.B. Jo-
hnson M.J. Lelevier, C.J. Schmitt, N.A. Wright & C.C.
Witt. 2011. Satellite imagery reveals new critical habitat
for endangered bird species in the high Andes of Peru.
Endangered Species Research 13: 145-157.
Bourg N.A., W.J. McShea & D.E. Gill. 2005. Putting the CART
before the search: successful habitat prediction for a rare
forest herb. Ecology 86: 2793-2804.
Buermann W., S. Saatchi, T.B. Smith, B.R. Zutta, J.A. Chaves, B.
Mil & C.H. Graham. 2008. Predicting species distribution
across the Amazonian and Andean regions using remote
sensing data. Journal of Biogeography 35: 1160-1176.
Cierjacks A., S. Salgado, K. Wesche & I. Hensen. 2008. Post-fre
population dynamics of two tree species in high-altitude Po-
lylepis forests of Central Ecuador. Biotropica 40: 176-182.
Costa G.C., C. Nogueira, R.B. Machado & G.R. Colli. 2010. Sam-
pling bias and the use of ecological niche modeling in
conservation planning: a feld evaluation in a biodiversity
hotspot. Biodiversity Conservation 19: 883-899.
Ellenberg H. 1979. Mans infuence on tropical mountain ecosystems
in South America. Journal of Ecology 67: 401-416.
Fjelds J. & M. Kessler. 1996. Conserving the biological diversity
of Polylepis forests of the highlands of Peru and Bolivia.
Nordic Foundation for Development and Ecology (NOR-
DECO), Copenhagen.
Fjelds, J. & M. Kessler. 2004. Conservacin de la biodiversidad de
los bosques de Polylepis de las tierras altas de Bolivia: Una
contribucin al manejo sustentable en los Andes. Centro
para la Investigacin de la Diversidad Cultural y Biolgica
de las Bosques Pluviales Andinos (DIVA), Dinamarca.
Fjelds J. 2002. Polylepis forests - vestiges of a vanishing ecosystem
in the Andes. Ecotropica 8: 111-123.
Garca-Nuez C., F. Rada, C. Boero, J. Gonzlez, M. Gallardo,
A. Azcar, M. Liberman-Cruz, M. Hilal & F. Prado.
2004. Leaf gas exchange and water relations in Polylepis
tarapacana at extreme altitudes in the Bolivian Andes.
Photosynthetica 42: 133-138.
Guisan A. & W. Thuiller. 2005. Predicting species distribution:
offering more than simple habitat models. Ecological
Letters 8: 993-1009
Hanselman J.A., W.D. Gosling, G.M. Paduano & M.B. Bush. 2005.
Contrasting pollen histories of MIS 5e and the Holocene
from Lake Titicaca (Bolivia/Peru). Journal of Quaternary
Science, 20: 663-670.
Hansen M.C., R.S. Defries, J.R.G. Townshend, R. Sohlberg, C.
Dimiceli & M. Carroll. 2002. Towards an operational
MODIS continuous feld of percent tree cover algorithm:
examples using AVHRR and MODIS data. Remote Sen-
sing of Environment 83: 303-319.
Hensen I., A. Cierjacks, H. Hirsch, M. Kessler, K. Romoleroux, D.
Renison & K. Wesche. 2012. Historic and recent fragmen-
tation coupled with altitude affect the genetic population
structure of one of the worlds highest tropical tree line
species. Global Ecology and Biogeography, 21: 455-464.
Hernandez P.A., I. Franke, S.K. Herzog, V. Pacheco, L. Paniagua,
H.L. Quintana, A. Soto, J.J. Swenson, C. Tovar, T.H.
Valqui, J. Vargas & B.E. Young. 2008. Predicting species
distributions in poorly studied landscapes. Biodiversity
Conservation 17: 1353-1366.
212
Zutta et al.
Rev. peru. biol. 19(2): 205 - 212 (Agosto 2012)
Hijmans R.J., S.E. Cameron, J.L. Parra, P.G. Jones & A. Jarvis.
2005. Very high resolution interpolated climate surface
for global land areas. International Journal of Climatology
25: 1965-1978.
IUCN. 2011. (en linea). 2011 International Union for Conservation
of Nature (IUCN) Red List of Threatened Species, <http://
www.iucnredlist.org>. Accesso 07/03/2012.
Jameson J.S. & P.M. Ramsay. 2007. Changes in high-altitude Polyle-
pis forest cover and quality in the Cordillera de Vilcanota,
Per, 1956-2005. Biological Conservation 138: 38-46.
Julio N.B., J.C. Rondan Dueas, D. Renison & I. Hensen. 2011.
Genetic structure and diversity of Polylepis australis
(Rosaceae) tree populations from central Argentina: impli-
cations for forest conservation. Silvae Genetica 60: 55-61.
Kessler M. & A.N. Schmidt-Lebuhn. 2006. Taxonomical and dis-
tributional notes on Polylepis (Rosaceae). Organisms,
Diversity & Evolution 6: 67-69.
Kessler M. 1995. The genus Polylepis (Rosaceae) in Bolivia. Can-
dollea 50: 131-171.
Kessler M. 2002. The Polylepis problem: Where do we stand. Eco-
tropica 8: 97-110.
Kunz L.B. & D.G. Long. 2005. Calibrating seawinds and QuikSCAT
scatterometers using natural land targets. IEEE Geoscience
and Remote Sensing Letters 2: 182-186.
Lloyd H. & S.J. Marsden. 2011. Between-patch bird movements
within a high-Andean Polylepis woodland/matrix lands-
cape: Implications for habitat restoration. Restoration
Ecology 19: 74-82.
Lloyd H. 2008. Abundance and patterns of rarity of Polylepis birds
in the Cordillera Vilcanota, southern Per: implications
for habitat management strategies. Bird Conservation
International 18: 164-180.
Mendoza W. & A. Cano. 2011. Diversidad del gnero Polylepis (Ro-
saceae, Sanguisorbeae) en los Andes peruanos: Diversity
of the genus Polylepis (Rosaceae, Sanguisorbeae) in the
Peruvian Andes. Revista Peruana de Biologa 18: 197-200.
Myneni R.B., S. Hoffman, Y. Knyazikhin, J.L. Privette, J. Glassy, Y.
Tian, Y. Wang, X. Song, Y. Zhang, G.R. Smith, A. Lotsch,
M. Friedl, J.T. Morisette, P. Votava, R.R. Nemani & S.W.
Running. 2002. Global products of vegetation leaf area
and fraction absorbed PAR from year one of MODIS data.
Remote Sensing of Environment 83: 214-231.
Philips S.J., R.P. Anderson & R.E. Schapire. 2006. Maximum entropy
modeling of species geographic distributions. Ecological
Modeling 190: 231-259.
Puhakka L., M. Salo & I.E. Sksjrvi. 2011. Bird diversity, bird-
watching tourism and conservation in Peru: a geographic
analysis. PLoS ONE 6: e26786.doi:10.1371/journal.
pone.0026786.
Purcell J. & A. Brelsford. 2004. Reassessing the causes of decline
of Polylepis, a tropical subalpine forest. Ecotropica 10:
155-158.
Purcell J., A. Brelsford & M. Kessler. 2004. The worlds highest
forest. American Scientist 92: 454-461.
Quinteros-Casaverde N., C.F. Flores-Negrn & D.A. Williams.
2012. Low genetic diversity and fragmentation effects in a
wind-pollinated tree, Polylepis multijuga Plige (Rosaceae)
in the high Andes. Conservation Genetics 13: 593-603.
Raxworthy C.J., E. Martinez-Meyer, N. Horning, R.A. Nussbaum,
G.E. Schneider, M.A. Ortega-Huerta & A.T. Peterson.
2003. Predicting distribution of known and unknown reptil
especies in Madagascar. Nature 426: 837-841.
Renison D., I. Hensen & R. Suarez. 2011. Landscape structural
complexity of high-mountain Polylepis australis forests:
a new aspect of restoration goals. Restoration Ecology
19: 390-398.
Riordan E.C. & P.W. Rundel. 2009. Modeling the distribution of a
threatened habitat: the California sage scrub. Journal of
Biogeography 36: 2176-2188.
Rios S.S, H. Lloyd & A. Valds-Velsquez. 2011. Bird species ri-
chness, diversity and abundante in Polylepis woodlands,
Huascaran biosphere reserve, Peru. Studies on Neotropical
Fauna and Environment 46: 69-76.
Rodriguez L.O. & K.R. Young. 2000. Biological diversity of Peru:
determining priority areas for conservation. AMBIO 29:
329-337.
Schmidt-Lebuhn A.N., M. Kessler & M. Kumar. 2006. Promiscuity
in the Andes: species relationships in Polylepis (Rosaceae,
Sanguisorbeae) based on AFLP and morphology. Syste-
matic Botany 31: 547-559.
Schmidt-Lebuhn A.N., M. Kumar & M. Kessler. 2006. An as-
sessment of the genetic population structure of two species
of Polylepis Ruiz & Pav. (Rosaceae) in the Chilean Andes.
Flora 201: 317-325.
Schmidt-Lebuhn A.N., P. Seltmann & M. Kessler. 2007. Consequen-
ces of the pollination system on genetic structure and pat-
tern of species distribution in the Andean genus Polylepis
(Rosaceae): a comparative study. Plant Systematics and
Evolution 266: 91-103.
Servat G.P., C.W. Mendoza & J.A. Ochoa C. 2002. Flora y fauna de
cuatro bosques de Polylepis (Rosaceae) en la Cordillera
del Vilcanota (Cusco, Peru). Ecologa Aplicada 1: 25-35.
Simpson B.B. 1986. Speciation and specialization of Polylepis in
the Andes. In: F. Vuillemeir and M. Monastrio, eds. High
Altitude Tropical Biogeography, Oxford University Press,
Oxford. Pp. 304-315.
Soutullo A., M. DeCastro & V. Urios. 2008. Linking political and
scientifcally derived targets for global biodiversity con-
servation: implications for the expansion of the global
network of protected areas. Diversity and Distribution
14:604-613.
Teich I., A.M. Cingolani, D. Renison, I. Hensen & M.A. Giorgis.
2005. Do domestic herbivores retard Polylepis australis
Bitt. woodland recovery in the mountains of Crdoba,
Argentina? Forest Ecology and Management 219: 229-241.
Thomassen H.A., T. Fuller, W. Buermann, B. Mil, C.M. Kieswetter,
P. Jarrn-V., S.E. Cameron, E. Mason, R. Schweizer, J.
Schlunegger, J. Chan, O. Wang, M. Peralvo, C.J. Schnei-
der, C.H. Graham, J.P. Pollinger, S. Saatchi, R.K. Wayne &
T.B. Smith. 2011. Mapping evolutionary process: a mutli-
taxa approach to conservation prioritization. Evolutionary
Applications 4: 397-413.
Wilson K.A., M.I. Westphal, H.P. Possingham & J. Elith. 2005. Sen-
sitivity of conservation planning to different apporoaches
to using predicted species distributions on the east slope
of the Andes in Peru and Bolivia. Biological Conservation
122: 99-112.
213
El nido de ScolomyS melanopS
Rev. peru. biol. 19(2): 213 - 216 (August 2012)
Rev. peru. biol. 19(2): 213 - 216 (Agosto 2012)
Facultad de Ciencias Biolgicas UNMSM
ISSN 1561-0837
Descripcin del nido de Scolomys melanops (Rodentia, Cricetidae) y
su relacin con Lepidocaryum tenue (Arecales, Arecaceae)
Edgardo M. Rengifo
1
y Rolando Aquino
2
Description of Scolomys melanops nest (Rodentia, Cricetidae) and its
relationship with Lepidocaryum tenue (Arecales, Arecaceae)
1 Museo de Historia Natural de la
Universidad Nacional Mayor de
San Marcos, Apartado 14-0434,
Lima-14, Per. Telfono: (511) 471
0117 anexo 31.
2 Instituto de Ciencias Biolgicas
Antonio Raimondi, Facultad de
Ciencias Biolgicas, Universidad
Nacional Mayor de San Marcos. P.
O. Box 575 Iquitos, Per.
Email Edgardo M. Rengifo:
edgar_mrv@hotmail.com
NOTA CIENTFICA
Presentado: 16/02/2012
Aceptado: 17/09/2012
Publicado online: 10/11/2012
Resumen
Se describe el nido de Scolomys melanops (Rodentia: Cricetidae), encontrado en el Nororiente de Per en un
hbitat de Colinas bajas. Se identifc tres estructuras en el nido: a) Orifcio de acceso y salida; b) Tnel y c)
Dormidero, en este ltimo se encontraron restos de frutos, fbras del pednculo y tallos de Lepidocaryum tenue
(Arecales: Arecaceae) Irapay. Se sugiere en base al material vegetal encontrado en el nido, la existencia de
una relacin de depredacin entre el roedor Scolomys melanops y la palmera Lepidocaryum tenue.
Palabras clave: Scolomys melanops; Lepidicaryun tenue; nido; depredacin.
Abstract
We describe the nest of Scolomys melanops, (Rodentia: Cricetidae) found in low hill habitats in northeastern
Peru. We identifed three structures in the nest: a) an orifce of access and exit; b) a tunnel and c) roosting area.
In the last structure, we found fruit and fber remains of peduncle and stems of Lepidocaryum tenue (Arecales:
Arecaceae) Irapay. Based on the vegetal remains found in the nest, we suggest the existence of a predation
relationship between the rodent Scolomys melanops and the palm Lepidocaryum tenue.
Keywords: Scolomys melanops; Lepidicarun tenue; nest; predation.
Scolomys (Rodentia: Cricetidae), es un gnero de roedor Sig-
modontino que ocurre en la cuenca amaznica, con distribucin
geogrfca en el oriente de Ecuador, sur de Colombia, nororiente
del Per y occidente de Brasil (Patton & da Silva 1995, Em-
mons & Feer 1997, Eisenberg & Redford 1999, Castro-Revelo
& Zapata-Rios 2001, Gomez-Laverde et al 2004), alcanza los
1150 m de altitud (Nowak 1991) y habita bosques de tierra frme
(Emmons & Feer 1997, Patton et al. 2000). El gnero est re-
presentado por dos especies segn Gomez-Laverde et al. (2004):
Scolomys ucayalensis, descrito por Pacheco (1991), en base a dos
especmenes de Jenaro Herrera al margen derecha del ro Uca-
yali en Per, y Scolomys melanops descrito por Anthony (1924)
en base a seis especmenes de la localidad de Mera en Ecuador.
Scolomys melanops es considerada una especie rara y muy poco
conocida (Hice 2001, Hice & Schmidly 2002), se distribuye al
norte de los ros Maran y Amazonas en bosques primarios no
perturbados con sotobosque denso (Emmons & Feer 1997). Esta
especie es bsicamente terrestre, aunque tambin fue encontra-
da a 1 m de altura sobre rboles cados (Hice 2001). Aspectos
de su reproduccin fueron reportados por Hice (2003), quien
encontr dos hembras preadas, una en marzo y otra en abril
y ocho machos con testculos escrotales en los meses de marzo,
octubre y noviembre. Otra informacin sobre la ecologa de la
especie no es conocida (Tirira 2007).
El presente trabajo contribuye con el conocimiento sobre
algunos aspectos de la ecologa y la historia natural de Scolomys
melanops, describiendo las estructuras y caractersticas del nido de
esta especie, as mismo, se menciona la relacin de depredacin
entre esta especie y la palmera cespitosa Irapay Lepidicaryun
tenue Mart. (Arecales, Arecaceae), en base a restos de fruto y fbras
del tallo encontradas dentro del nido. Los datos presentados sern
tiles para la elaboracin de planes de conservacin destinada a
proteger ambas especies.
Durante un inventario llevado a cabo en los bosques de la
cuenca alta del Rio Itaya, el 30 de julio de 2008, en el campamen-
to Yanayacu (41348S, 734210W), ubicado en el distrito
de San Juan Bautista, Provincia de Maynas, Departamento de
Loreto, en el nororiente de Per, a 70 km al suroeste de Iquitos
(Fig. 1), fue encontrado el nido de un individuo de Scolomys
Figura 1. Mapa de Ubicacin geogrfca de la localidad Yanayacu
(estrella) en Loreto Per, donde fue registrado el nido de Scolomys
malanops.
214
Rengifo & Aquino
Rev. peru. biol. 19(2): 213 - 216 (Agosto 2012)
melanops. El hbitat corresponda a bosques de colinas bajas
(Encarnacin 1985); las colinas variaban entre 10 a 20% de
pendientes, con rboles robustos de 25 m de altura y algunos
emergentes que alcanzaban los 30 m; el sotobosque era denso,
suelo de tipo arcilloso cubierto de hojarascas de 2 a 3 cm de
espesor. Entre las especies vegetales dominantes del entorno del
nido fueron registradas Iriartea deltoidea Huacrapona, Calathea
lutea Bijao, Geonoma sp. Ponilla, Tachigali sp. Tangarana,
Minquartia guianensis Huacapu, Cedrelinga catenaeformis Tor-
nillo, Virola sp. Cuamala, Iryanthera sp. Cumala colorada y
Lepidocaryum tenue Irapay como especie dominante del rea,
la que cubra gran parte del sotobosque.
El hallazgo del nido fue accidental mientras se realizaba corte
de lea prximo al campamento, un individuo de Scolomys
melanops fue observado saliendo y entrando de dicho nido,
advirtiendo su lugar de nidacin, cabe resaltar que este indivi-
duo era el nico usuario de este nido. El individuo capturado
manualmente y luego liberado, corresponda a una hembra
adulta, con vagina no perforada, lo cual indicaba que no se
encontraba en periodo reproductivo. La identifcacin como S.
melanops se logr siguiendo las descripciones de Anthony (1924),
Eisenberg y Redford (1999), Emmons y Feer (1997) y Pacheco
(1991), quienes resaltan algunas caractersticas fenotpicas que
lo diferencian de otras especies presentes en el rea de estudio;
entre los principales caracteres observados tenemos: dorso de
color gris oscuro, pelaje compuesto por espinas entremezcladas
con pelos no modifcados, regin ventral gris, oreja corta, cola
desnuda y corta (menos de la mitad de la longitud total) y patas
posteriores anchas con el pulgar sin garra.
El nido fue encontrado entre las ramas de un rbol cado
en proceso descomposicin, lejos de cuerpos de agua, siendo
el ms prximo una pequea quebrada de 2 m de cauce loca-
lizada a una distancia aproximada de 50 m. En la descripcin
del nido se consideraron las medidas del dimetro del ancho,
largo y alto de las estructuras encontradas, adems se anotaron
las caractersticas ms importantes y especies vegetales con las
que estuvo construido el nido. En el nido fueron diferenciadas
tres estructuras: a) Orifcio de acceso y salida, b) Tnel y c)
Dormidero (Fig. 2).
a) Orifcio de acceso y salida: Ubicado entre dos ramas
de 12 y 7 cm de dimetro respectivamente y estratgicamente
construido para evitar la entrada de agua de lluvias hacia el
interior, pues la entrada se encontraba formando un ngulo
de 85 en relacin al suelo, sobresaliendo la parte superior, lo
cual evitara la escorrenta hacia el interior . Esta estructura
constituye la parte ms visible del nido, cuyo orifcio de 2,5 cm
de dimetro se encontraba libre de obstculos que difculten el
acceso hacia el nido.
b) Tnel: Seguido del orifcio de acceso y salida, con 7 cm
de longitud; este tnel conectaba la entrada del nido con el
dormidero; se caracteriz por ser de forma cilndrica, angosta
cerca a la entrada y progresivamente ensanchada hacia el dor-
midero, se caracteriz por estar recubierto de las raicillas del
entono del suelo.
c) Dormidero: Localizado inmediatamente despus del
extremo posterior del tnel; tiene forma ovoide, las medidas
fueron de 6,5 cm de largo (desde la boca del tnel hasta la pared
Figura 2. Esquema representativo del nido de Scolomys melanops: a) Orifcio de acceso y salida, b) Tnel y c) Dormidero; la fecha seala
el acceso al nido.
215
El nido de ScolomyS melanopS
Rev. peru. biol. 19(2): 213 - 216 (August 2012)
posterior), 6 cm de ancho (entre las paredes laterales) y 6 cm de
alto (del piso hasta la pared superior). El dormidero corresponde
a la parte ms seca del nido, destac entre sus componentes
un montculo en forma de colchn, que sera usado para el
descanso y construido con fbra del tallo de Lepidocaryum tenue
Irapay; adems, se encontr el endocarpio del fruto de esta
especie vegetal, con rasgos de haber sido mordido y usado como
alimento por el individuo capturado.
El tipo de hbitat donde fue encontrado el nido, bosques
de colina bajas, ofrece caractersticas fsiogrfcas que evitan
que las someras y fugaces inundaciones producidas por las
lluvias afecte los lugares de nidacin de Scolomys melanops. La
preferencias del uso de estos hbitats es explicado por la lejana
del nido respecto a los cuerpos de agua. Esta interpretacin es
confrmada con los resultados obtenidos durante el inventario
de mamferos pequeos llevado a cabo en la cuenca alta del Rio
Itaya por Caldern y Rengifo (2010), ya que todas las capturas
de esta especie en ese estudio (n=7) fueron realizadas en Bosques
colinosos donde tambin se observ la abundancia de Lepido-
caryum tenue Irapay.
Tirira (2007) afrma el escaso conocimiento sobre ecologa
de Scolomys melanops. En base a la evidencia del material vegetal
encontrada en el nido, nos permite sostener que los frutos de
Lepidocaryum tenue constituyen una fuente alimenticia de este
roedor; asimismo, las fbras que forman el colchn del nido
ofrecen una forma de aislamiento del suelo hmedo. Una es-
tructura craneal especializada como arcos zigomticos anchos
(Anthony 1924) con incisivos proodontos (Pacheco 1991)
ayuda a S. melanops a desfbrar el tallo y pednculo de L. tenue,
adems le permite triturar fcilmente el pericarpio de un fruto
tan duro, como el que caracteriza a esta especie vegetal, el cual
est cubierto de escamas imbricadas (Uhl & Dransfeld 1987)
que hacen difcil el acceso al mesocarpio.
Existe una relacin de depredacin entre Scolomys melanops
(Rodentia: Cricetidae) y Lepidocaryum tenue (Arecales: Areca-
ceae); en base a estas observaciones inferimos que S. melanops
estara haciendo uso del fruto, pednculo y tallo tanto para
alimentacin como para la construccin de su nido.
Desafortunadamente la demanda de la hoja de Lepidocaryum
tenue Irapay es alta en muchas de las ciudades Amaznicas
(Mejia & Kahn 1996, Baslev et al. 2008, Navarro et al. 2011),
por lo que reas como la cuenca del ro Itaya son fuertemente
perturbadas por el corte de las hojas sin control ni planes de
manejo sobre esta palmera (Aquino et al. 2008, Aquino et al.
2009), modalidad de extraccin que est afectando a muchas
poblaciones de fauna silvestre, afectando redes trfcas y ame-
nazando las poblaciones que de forma directa o indirecta usan
este recurso, entre ellas se a Scolomys melanops, considerada una
especie rara en la amazonia (Hice 2001). Un manejo adecuado de
Lepidocaryum tenue ayudara por un lado a satisfacer su demanda
y por el otro evitara que muchas especies de fauna silvestre sean
afectadas por la extraccin de las hojas de esta palmera.
Agradecimientos
Al Centro Amaznico de Educacin Ambiental e Investiga-
cin (ACEER) a travs de su representante Aura Murrieta Torres
por el fnanciamiento de la investigacin, a Wendy Caldern
por su apoyo en la realizacin del trabajo de campo. Nuestro
reconocimiento a los guas: Gilmer Montero, Humberto Pea
(Q.E.P.D) y Hugo Pea, por el gran esfuerzo realizado durante
las actividades de campo. A Filomeno Encarnacin, Vctor
Pacheco, Mara Peralta y Natali Hurtado por la revisin previa
y comentarios sobre el manuscrito.
Literatura citada
Anthony H.E. 1924. Preliminary report on Ecuadorean mammals
No 6. Americam Museum Novitates 139: 1-9.
Aquino R., D. Gil & E. Pezo. 2009. Aspectos ecolgicos y sos-
tenibilidad de la caza del majs (Cuniculus paca) en la
cuenca del ro Itaya, Amazona peruana. Revista Peruana
de Biologa 16(1): 067- 072.
Aquino R., W. Terrones, F. Cornejo & E.W. Heymann. 2008. Geo-
graphic distribution and possible taxonomic distinction of
Callicebus torquatus populations (Pitheciidae: Primates)
in Peruvian Amazonia. American Journal of Primatology
70: 11811186.
Balslev H., C. Grandez, N.Y. Paniagua-Zambrana, et. al. 2008.
Palmas (Arecaceae) tiles en los alrededores de Iquitos,
Amazona Peruana. Revista Peruana de Biologa. 15(1):
121-132.
Caldern W. & E.M. Rengifo. 2010. Diversidad y uso de Hbitat
por micromamferos en la cuenca alta del rio Itaya. Tesis,
Bilogo. Facultad de Ciencias Biolgicas Universidad
Nacional de la Amazonia Peruana.
Castro-Revelo I. & G.A. Zpata-Ros. 2001. First account and geo-
graphic range extension for Scolomys juruaense (Roden-
tia: Muridae) from eastern Ecuador. Revista de Biologa
Tropical 49: 769.
Eisenberg J.F. & K.H. Redford. 1999. Mammals of the Neotropics.
Volume 3. The Central Neotropics: Ecuador, Peru, Bolivia,
Brazil. University of Chicago Press, Chicago. 609 pp.
Gomez-Laverde M., R.P. Anderson & L.F. Garcia. 2004. Integrated
systematic reevaluation of the Amazonian genus Sco-
lomys (Rodentia: Sigmodontinae). Mammalian Biology
69:119139.
Hice C.L. & D.J. Schmidly. 2002. The effectiveness of pitfall traps
for sampling small mammals in the Amazon basin. Mas-
tozoologia Neotropical 9(1): 85-89.
Hice C.L. 2001. Record of a few rare mammals from northeastern
Peru. Mammalian Biology 66:317-319.
Hice C.L. 2003. The Non-Volant Mammals of the Estacin Biolgica
Allpahuayo: Assessment of the Natural History and Com-
munity Ecology of a Proposed Reserve. Ph.D. dissertation
Texas Tech University, Texas, EEUU.
Kahn F. & F. Moussa. 1994. Las palmeras del Per - Colecciones,
Patrones de distribucin geogrfca, ecologa, estatutos
de conservacin, nombres vernculos, utilizaciones.
IFEA, Lima.
Mejia K. & F. Kahn. 1996. Biologa, ecologa y utilizacin del
Irapay (Lepidocaryum gracile Martius). Folia Amaznica
8(1): 19-28.
Navarro J.A., G. Galeano & R. Bernal. 2011. Impact of leaf harvest
on populations of Lepidocaryum tenue, an Amazonian
understory palm used for thatching. Tropical Conservation
Science 4(1):25-38.
Nowak R. 1991. Walker's Mammals of the World. 5th edition, Vol.
I. Johns Hopkins University Press, Baltimore and London.
642 pp.
Pacheco V. 1991. A new species of Scolomys (Muridae: Sigmodon-
tinae) from Peru. Publicaciones del Museo de Historia
Natural Javier Prado, UNMSM, Serie A 37:13.
Patton J. L. & M.N.F. Da Silva. 1995. A review of the spiny mouse
genus Scolomys (Rodentia: Muridae: Sigmodontinae)
with the description of a new species from the western
Amazon of Brazil. Proceedings of the Biological of Society
Washington 108: 319-337.
216
Rengifo & Aquino
Rev. peru. biol. 19(2): 213 - 216 (Agosto 2012)
Patton J.L, M.N.F. Da Silva & J.R. Malcolm. 2000. Mammals of the
Rio Juru and the evolutionary and ecological diversifca-
tion of Amazonia. Bulletin of the American Museum of
Natural History 244: 1-306.
Tirira, D. 2007. Gua de Campo de los Mamferos del Ecuador.
Ediciones Murcilago Blanco. Publicacin especial sobre
los mamferos del Ecuador, Quito, Ecuador.
Uhl N.W. & J. Dransfeld. 1987. Genera Palmarum. A Classifcation
of Palms based on work of Harold E. Moore, Jr. Allen
Press, Lawrence, Kansas. 610 pp.
217
Ampliacin de la distribucin de SephanoiDeS SephanioDeS
Rev. peru. biol. 19(2): 217 - 218 (August 2012)
Rev. peru. biol. 19(2): 217 - 218 (Agosto 2012)
Facultad de Ciencias Biolgicas UNMSM
ISSN 1561-0837
Ampliacin de la distribucin de Sephanoides sephaniodes (Trochilidae:
Aves) en Argentina
Nancy Vernica Marinero
1
, Ricardo Omar Cortez
2
, Eduardo Alfredo Sanabria
2
*,
Lorena Beatriz Quiroga
2
Expansion of the distribution of Sephanoides sephaniodes (Trochilidae:
Aves) in Argentina
1 Instituto de Diversidad y Eco-
loga Animal, Centro de Zoologa
Aplicada, Universidad Nacional de
Crdoba, Argentina. Rondeu 798,
Nueva Crdoba. Crdoba (CP
5000). Argentina
2 Departamento de Biologa, Uni-
versidad Nacional de San Juan,
Argentina.
Email Nancy Vernica Marinero:
veronicamarinero@gmail.com
Email Ricardo Omar Cortez:
cortezricard@gmail.com
Email Eduardo Alfredo Sanabria:
sanabria.eduardoa@gmail.com
Email Lorena Beatriz Quiroga:
quirogalb@gmail.com
NOTA CIENTFICA
Presentado: 03/04/2012
Aceptado: 29/08/2012
Publicado online: 10/11/2012
Resumen
El presente trabajo reporta la ampliacin de la distribucin de Sephanoides sephaniodes hacia el centro-oeste
de Argentina. Se analizan posibles causas de la dispersin y consecuencias ecolgicas de la misma.
Palabras clave: Aves, Distribucin, Sephanoides sephaniodes, San Juan, Argentina.
Abstract
The present work report the expansion of the distribution of Sephanoides sephaniodes toward the west-central
of Argentina. It discusses possible causes of dispersion and ecological consequences of them.
Keyword: Birds, Distribution, Sephanoides sephaniodes, San Juan, Argentina.
La distribucin del Picafor Rub Sephanoides sephaniodes
(Lesson 1826) se encuentra restringida hacia el sur del continente
Americano, abarcando los pases de Chile y Argentina (Olrog
1984). Especfcamente, en Argentina se distribuye desde Tierra
del Fuego a Mendoza (Narosky & Izurieta 2003) (Fig. 1). Habita
los bosques andino-patagnicos representados por Araucaria sp.
(Mata et al. 2006) y Nothofagus sp. (Fraga et al. 1997), aunque
tambin puede observarse en reas de estepa (Grigera & Trejo
2009). El Picafor Rub se identifca por la presencia de corona
rojo brillante, la zona ventral es griscea y en la garganta exhibe
notables pecas verdosas (De la Pea & Rumboll 1998; Narozky
& Zurieta 2003; Mata et al. 2006).
Actualmente, esta especie exhibe registros ocasionales en
latitudes menores a las habituales. Hacia el este de Argentina, se
detectaron individuos en la provincia de Buenos Aires (Maugeri
2002) y Santa F (Giarduz 2008) mientras que, hacia el oeste hay
dos registros en el sur de la provincia de San Juan (Lucero 2009).
El 23 de Setiembre del 2010, se realiz un nuevo avistamiento
de S. sephaniodes en la Sierra de la Dehesa en la provincia de San
Juan (312988,7S 689170,6W), constituyendo el registro
ms septentrional para la especie hacia el centro-oeste de Argen-
tina. El individuo avistado era un macho adulto que pernoctaba
en una grieta longitudinal de una pared rocosa, contigua a una
naciente de agua (Fig. 2). El ambiente pertenece a la ecorregin
del Monte y a diferencia de los bosques andino-patagnicos,
se caracterizada por su intensa aridez. Las precipitaciones son
estivales, menores a los 200 mm anuales en tanto que, la tem-
peratura media anual es de 24 C (Poblete & Minetti 1999).
Fisonmicamente, el Monte es un mosaico representado princi-
palmente por una estepa arbustiva de Larrea spp., acompaada
de halftos como Atriplex sp., siendo escaso el estrato arbreo
representado por Prosopis spp. (Cabrera 1976).
El Picafor Rub puede cambiar de estrategias comportamen-
tales de forrajeo, dependiendo de las condiciones de termorregu-
lacin y costos de la alimentacin que deba enfrentar (Fernndez
et al. 2002). Adems, puede evitar la prdida de agua corporal ante
la escasez de la misma en el ambiente (Bakken & Sabbat 2006).
Teniendo en cuenta por un lado, la tolerancia de esta especie ante
diferentes escenarios ecolgicos y por otro, los registros recientes
hacia el norte de Argentina, es probable que durante los meses ms
fros S. sephaniodes se comporte como migrante estacional. Este es-
tado ha sido sugerido para el centro-este de Argentina hacia donde
se desplazaran pequeas poblaciones durante otoo (Olrog 1979).
El desplazamiento de individuos implicara que superpongan su
rango de distribucin con el de otros picafores. Particularmente,
Figura 1. Distribucin de Sephanoides sephaniodes en Argentina,
reas de posible observacin y lugar donde se realiz el nuevo
registro en el centro-oeste de Argentina (Modifcado desde Narosky
& Izurieta 2003).
218
Marinero et al.
Rev. peru. biol. 19(2): 217 - 218 (Agosto 2012)
en la Sierra de la Dehesa los individuos solaparan su nicho con
el Picafor Andino (Oreotrochilus leucopleurus), Picafor Gigante
(Patagona gigas) y Picafor Cometa (Sappho sparganura) (Olrog
1986; Narozky & Zurieta 2003; Mata & Rumboll 2006). Efec-
tivamente, Lucero (2009) reporta disputas entre el Picafor Rub
y el Picafor Andino por fores de Nicotiana glauca, demostrando
competencia por los recursos alimenticios, que en los ambientes
ridos y semiridos son escasos.
De esta manera, en el contexto de las evidencias fsiolgicas
sobre las estrategias y tolerancia de esta especie, predecimos que
continuarn surgiendo nuevos registros de S. sephaniodes en
ambientes ridos y/o paisajsticamente diferentes a los bosques
andino-patagnicos donde la especie es residente.
Literatura citada
Bakken H. & P. Sabbat. 2006. Gastrointestinal and renal responses to
water the green-backed frecrown (Sephanoides sephanoi-
des), a South American hummingbird. American Journal
of physiology Regulatory, Integrative and comparative
physiology 291: 830-836.
Cabrera A. 1976. Regiones ftogeogrfcas Argentinas. Enciclope-
dia de agricultura y jardinera. Tomo II. ACME. Buenos
Aires. Argentina.
De la Pea M. & M. Rumboll. 1998. Birds of southern South America
and Antarctica. Priceton. New Jersey.
Fraga M., E. Ruffni & D. Grigera. 1997. Interacciones entre el
picaflor rub Sephanoides sephaniodes y plantas del
bosque subantrtico en el parque nacional Nahuel Huapi,
Argentina. Hornero 14: 224-234.
Fernndez M., M. Lpez-Calleja & F. Bozinovic. 2002. Interplay
between the energetics of foraging and thermoregulatory
costs in the green-backed frecrown hummingbird Sep-
hanoides sephaniodes. Journal of Zoology 258: 319-326.
Giarduz C. 2008. Picafor cometa (Sappho sparganura), Picafor
rub (Sephanoides sephaniodes) y Picafor garganta blanca
(Leucochloris albicollis) en Rosario, provincia de Santa
F, Argentina. Nuestras Aves 53: 31-33.
Grigera D. & A. Trejo. 2009. Aves continentales de las provincias
del Neuqun, Ro Negro y Chubut. Distribucin y estado
de conservacin. Secretara de Investigacin del Centro
Regional Universitario Bariloche. Universidad Nacional
del Comahue.
Lucero F. 2009. Aves nuevas, raras o con pocos registros para las pro-
vincias de Mendoza y San Juan. Nuestras aves 54: 57-62.
Mata J., F. Erize & M. Rumboll. 2006. Aves de Sudamrica: Gua de
campo Collins Primera edicin. Letemendia. Buenos Aires.
Maugeri F. 2002. Aves nuevas o poco conocidas para el sudoeste
de la provincia de Buenos Aires, Argentina. Nuestras aves
44: 16-19.
Narosky T. & D. Izurieta. 2003. Gua para la identifcacin de las
aves de Argentina y Uruguay. Asociacin Ornitolgica
del Plata. 340pp.
Olrog C. 1979. Nueva lista de la avifauna Argentina. Opera Lilloana
27:1-324.
Olrog C. 1984. Las aves argentinas. Una nueva gua de campo.
Administracin de Parques Nacionales. 352 pp.
Poblete G. & J. Minetti. 1999. Confguracin espacial del clima de
San Juan. Sntesis del cuaternario de la provincia de San
Juan. INGEO. UNSJ. San Juan.
Figura 2. A) Pared rocosa don-
de fue detectado Sephanoides
sephaniodes. B) Posicin en
que pernoctaba el individuo
en la grieta. C) Visualizacin
de caracteres diagnsticos:
corona rojo brillante y pecas
en la garganta. Fotos: Ricardo
Cortez y Eduardo Sanabria.
* Actual lugar de trabajo: Laboratorio de Investigaciones Androlgicas de Men-
doza. Instituto de Histologa y Embriologa de Mendoza, Facultad de Ciencias
Mdicas, Universidad Nacional de Cuyo. Centro de Investigaciones en Ciencia
y Tcnica de Mendoza, CONICET, Mendoza, Argentina.
219
Herbarium Areqvipense (HUSA)
Rev. peru. biol. 19(2): 219 - 222 (August 2012)
Rev. peru. biol. 19(2): 219 - 222 (Agosto 2012)
Facultad de Ciencias Biolgicas UNMSM
ISSN 1561-0837
Herbarium Areqvipense (HUSA): informatizacin y representatividad
de su coleccin
Italo F. Trevio
1,2,
*, Diego A. Sotomayor
1,2,3
, Marco A. Cueva
1,2
, Rafael Perez
1,2
,
Laura Cceres
1,2
, Daniel Ramos
1,2
, Edgardo M. Ortiz
1,2,4
, Victor Quipuscoa
1,2
Herbarium Areqvipense (HUSA): computerization and representativeness
of its collection
1 Herbarium Areqvipense (HUSA),
Universidad Nacional de San Agus-
tn de Arequipa, Daniel Alcides
Carrin s/n
2 Grupo de Investigacin de la
Diversidad Biolgica del sur del
Per (DIBIOS)
3 Department of Geography, York
University, 4700 Keele Street,
Toronto, M3J 1P3, ON, Canada
4 Section of Integrative Biology and
the Plant Resources Center, The
University of Texas at Austin, Texas
78712 U.S.A.
* Autor para correspondencia
Email Italo Trevio:
ifrant01@gmail.com
NOTA CIENTFICA
Presentado: 18/05/2012
Aceptado: 16/10/2012
Publicado online: 10/11/2012
Resumen
Las colecciones cientfcas y entre ellas los herbarios representan importantes fuentes de informacin y
enseanza para los investigadores y profesionales en ciencias biolgicas. El Herbarium Areqvipense (HUSA),
registrado en el Index Herbariorum desde el ao 2004, alberga una de las colecciones ms importantes del
Per. En este trabajo brindamos informacin de la coleccin y damos a conocer su representatividad para la
fora peruana. El HUSA cuenta con ms de 11000 especmenes registrados, distribuidos en ms de 2300 espe-
cies, correspondientes en su mayor parte a Magnoliophyta y Pteridophyta (ca. 98%), y en menor proporcin
por Basidiomycetes y Ascomycetes (Hongos y Lquenes) y Bryophyta (Musgos). Los especmenes provienen
de 23 departamentos del Per, donde Arequipa presenta el mayor nmero de individuos colectados (3375),
equivalente al 31% de la coleccin. Las familias Asteraceae y de Solanaceae son las mejor representadas
con 1571 y 964 especmenes, respectivamente. La mayor cantidad de especmenes georeferenciados corre-
sponden a la zona de vida Bosque muy hmedo bajo tropical con el 15%, seguido por el Bosque muy hmedo
premontano tropical con el 8%. Se dan adems a conocer los tipos presentes en la coleccin, as como una
resea del desarrollo del HUSA desde su creacin.
Palabras clave: coleccin cientfca, herbario, HUSA, tipos, fora peruana.
Abstract
Scientifc collections and herbaria are essential sources of information and education for researchers and
practitioners in biological sciences. The Herbarium Areqvipense (HUSA), registered at Index Herbariorum
since 2004, holds one of the most important collections in Peru. In this paper we provide information about the
collection, and its representativeness for the Peruvian fora. HUSA has more than 11000 specimens recorded
to date, with more than 2300 determined species, consisting mostly of Magnoliophyta and Pteridophyta (ca.
98%), and a smaller proportion of Basidiomycetes, Ascomycetes (fungi and lichens) and Bryophyta (mosses).
The collection includes specimens from 23 departments of Peru, where the samples belonging to Arequipa
have the largest number of individuals collected (3375) accounting for 31% of the collection. Asteraceae and
Solanaceae are the most collected with 1571 and 964 specimens, respectively. The majority of geo-referenced
specimens came from the tropical wet forest with 15%, followed by the tropical pre-montane wet forest with
8%. We also provide a list of the nomenclatural types and a brief summary of the history and development of
HUSA since its creation.
Keywords: collection, herbarium, HUSA, types, Peruvian fora.
Introduccin
Los herbarios representan importantes fuentes de informacin
sobre la diversidad de un territorio, as como para la investigacin
y docencia en ciencias biolgicas (Rao 1995). Las colecciones que
albergan los herbarios permiten la determinacin y la descripcin
de nuevas especies (Bebber et al. 2010); la informacin contenida
en sus voucher permiten conocer y aplicar modelos de distribu-
cin de especies (Loiselle et al. 2008), asimismo permite inferir
la distribucin histrica y potencial y fenologa de las especies
que albergan (Feeley et al. 2010, Montserrat-Marti et al. 1997),
y tambin es reconocida su importancia en la conservacin y
toma de decisiones (Rivers et al. 2011, Schatz 2002, Ponder
et al. 2001). Los herbarios permiten evaluar vacos en las colec-
ciones dentro de un territorio, establecer zonas prioritarias de
conservacin y determinar tendencias histricas en la presencia
de ciertas especies de inters (Squeo et al. 2008). Adems, los
herbarios juegan un rol educativo en la formacin de los futuros
profesionales en ciencias biolgicas.
La necesidad de documentar la biodiversidad toma ms im-
portancia en pases megadiversos como el Per, especialmente
cuando el crecimiento de la poblacin, la contaminacin, el
cambio climtico global y los requerimientos de mayores can-
tidades de recursos amenazan con el deterioro y prdida de los
hbitats, especies y biodiversidad en general, poniendo en peligro
la existencia de la humanidad. En este artculo, presentamos la
coleccin del Herbarium Areqvipense (HUSA), perteneciente a
la Universidad Nacional de San Agustn de Arequipa (UNSA).
Se incluye una breve resea histrica, la descripcin del proceso
de informatizacin de la coleccin, y fnalmente, el anlisis de
su representatividad para la fora peruana.
Desarrollo y actualidad
El Herbarium Areqvipense (HUSA) es una institucin
reconocida formalmente ante el Index Herbariorum (Holmgren
& Holmgren 2005) desde el ao 2004 y es parte de la Asociacin
de Herbarios del Per desde el ao 2002. Inici sus actividades de
220
Trevio et al.
Rev. peru. biol. 19(2): 219 - 222 (Agosto 2012)
formacin en la dcada de los 90s con la participacin e iniciativa
de los docentes del rea de Botnica de la Escuela Profesional
y Acadmica de Biologa, de la Universidad Nacional de San
Agustn de Arequipa (UNSA). Su inauguracin se realiz el 28
de abril del 2000 durante el VIII Congreso Nacional de Botnica
en Arequipa, teniendo como local desde la fecha el Aula 302 del
tercer piso de la Escuela Profesional y Acadmica de Biologa,
constituyndose en un principio como una herramienta de apoyo
para la formacin de los estudiantes en el campo de la Botnica.
Desde ese mismo ao el HUSA se asoci con el Grupo de
Investigacin de la Diversidad Biolgica del Sur (DIBIOS), grupo
en ese entonces conformado por estudiantes de aos superiores de
la Carrera Profesional de Biologa. Este grupo bajo la tutela del
profesor Blgo. Vctor Quipuscoa Silvestre (Docente del rea de
Botnica y subdirector del Herbario) particip activamente en el
manejo e implementacin del Herbario. Las muestras de perio-
dos previos que se encontraban almacenadas en cajas y pequeos
estantes fueron clasifcadas, procesadas e ingresadas a la coleccin,
descartndose aquellas que no contaban con datos de colecta y/o
caractersticas necesarias para su incorporacin al herbario.
En aos posteriores y mediante el esfuerzo de integrantes del
DIBIOS se organizaron salidas de campo para recolectar la fora
local con el propsito de incrementar la coleccin y teniendo
como destino distintas zonas de Arequipa y algunas veces otros
departamentos del sur del pas. Durante este proceso de creci-
miento de la coleccin, se propici la formacin e integracin de
ms estudiantes por medio del grupo DIBIOS para el apoyo en
el mantenimiento de la coleccin cientfca e ingreso de muestras,
arduo trabajo que permiti la indexacin del herbario en el ao
2004. Durante este proceso se destaca el apoyo del Dr. Michael
O. Dillon del Field Museum de Chicago, USA, quien es uno
de los principales benefactores e investigadores del Herbario y
que visit el Herbario desde sus inicios.
El rpido crecimiento de la coleccin, motiv la necesidad de
implementar herramientas que permitan un mejor manejo de la
coleccin. De esa manera, ese mismo ao 2004 se implement
la primera base de datos del Herbario, denominada Huthia y que
fue confeccionada por Edgardo M. Ortiz (miembro del grupo
DIBIOS) y desarrollada bajo la plataforma Microsoft Access XP
corriendo sobre el sistema operativo Windows 98 SE. La tabla
general que contiene los datos se elabor en base a la tabla de
especmenes para el Norte Peruano (DETBASE) desarrollada por
Nancy Hensold, una especialista en el Manejo de Colecciones
Tropicales del Field Museum de Chicago. La creacin de esta base
de datos permiti la re-estructuracin de la modalidad de ingreso
y ordenamiento de las muestras, iniciando la digitalizacin de los
datos de cada espcimen. Entre los aos 2005 y 2009 el Herbario
logr propiciar el desarrollo de proyectos, tesis, voluntariados;
fortaleciendo la formacin y desarrollo de los alumnos de Biologa,
as como el crecimiento de la coleccin. Entre 2010 y 2011, gracias
al fnanciamiento de la Red Interamericana de Informacin de
Biodiversidad (IABIN), se logr apoyo econmico para el ingreso
de ms de 3000 especmenes, as como su revisin y digitalizacin
de la coleccin entera.
En la actualidad el HUSA es una coleccin cientfca de ref-
erencia en el sur del Per y en especial para el departamento de
Arequipa. A pesar de tratarse de un Herbario joven, este se ha
convertido en un centro de investigacin y formacin de inves-
tigadores en el campo de la Botnica. Se practica un adecuado
manejo de la coleccin mantenindola en ptimo estado de
conservacin y organizacin. Las colecciones son utilizadas en
investigaciones taxonmicas, ecolgicas y de botnica econmica
entre otros, donde participan especialmente docentes de la UNSA,
investigadores de otras universidades peruanas y extranjeras, as
como alumnos y egresados de la Escuela de Biologa de la UNSA.
Desde su formacin, el HUSA tiene como misin la de contribuir
al desarrollo de nuestra sociedad y est abierto a la atencin y
servicio de la comunidad cientfca y pblico en general, siendo
las actividades ms comunes las determinaciones de especies y
realizacin de charlas a estudiantes de colegios. El herbario se
encuentra ubicado en el rea de Biomdicas de la Universidad
Nacional de San Agustn, avenida Daniel Alcides Carrin.
Informatizacin de la coleccin
Los datos del HUSA fueron informatizados como parte del
proyecto IABIN. La Red Interamericana de Informacin sobre
Biodiversidad (IABIN) fue creada con el objetivo de propiciar
la colaboracin tcnica y la coordinacin entre los pases de las
Amricas para recolectar, compartir y utilizar informacin sobre
biodiversidad que sea relevante para los procesos de toma de de-
cisiones, as como para la educacin en la regin, particularmente
para asuntos en la interface de desarrollo humano y conservacin
de la biodiversidad. La informacin de especmenes se registr
en la base de datos Huthia (versin 2.0). El protocolo seguido
para su desarrollo fue una modifcacin del trabajo de Martn-
Consuegra et al. (2009), el que consta de los siguientes pasos:
Digitalizacin y verifcacin de datos. Se digitalizaron los
datos de las boletas de los especmenes ingresados, en la versin
modifcada de Huthia, segn los estndares de DarwinCore.
Terminado este proceso la base de datos fue corroborada tres
veces con las boletas de los especmenes.
Revisin de la nomenclatura. Se realiz una actualizacin de
nomenclatura en la base de datos. Se adopt el sistema de clasif-
cacin de Lumbsch y Huhndorf (2007) para Lecanoromycetes y
el sistema de clasifcacin de Cronquist (1981) para Liliopsida
y Magnoliopsida, este ltimo con algunas modifcaciones en
ciertos grupos donde se adopt la clasifcacin utilizada por
Reveal y Chase (2011).
Geo-referenciacin de localidades. Con la asistencia del
programa ArcGIS v9.3 se realiz la asignacin de coordenadas
a especmenes que no presentaban datos de coordenadas en las
boletas. Para esto se tom en consideracin datos proporcio-
nados por los colectores a travs de consulta directa, bsqueda
de bibliografa relacionada a rangos de distribucin, hbitats y
reportes para la especie en la localidad de referencia. Los datos
fueron validados mediante un anlisis de superposicin de capas
de los lmites administrativos (localidad, distrito, provincia y de-
partamentos), zonas de vida, red hidrogrfca, red de carreteras,
centros poblados, elevacin, entre otros.
Alojamiento de datos integrados en la red IABIN. La base
de datos generada fue revisada dos veces ms, en bsqueda de
posibles errores, y posteriormente subida al servidor en el que se
alojan los datos de IABIN con la fnalidad de hacerla disponible
al pblico en general.
Riqueza de la coleccin
La coleccin del Herbario HUSA cuenta actualmente con
11251 especmenes correspondientes a ms de 2300 especies,
221
Herbarium Areqvipense (HUSA)
Rev. peru. biol. 19(2): 219 - 222 (August 2012)
74,67% de los cuales se encuentran geo-referenciados. Las
familias mejor representadas corresponden a Asteraceae (1571
especmenes), Solanaceae (964 especmenes), Ericaceae (467
especmenes), Fabaceae (460 especmenes) y Malvaceae (330
especmenes), teniendo las dems familias restantes por debajo
de 300 pliegos cada una. Los gneros con mayor nmero de
especmenes corresponden a Solanum (365), Nolana (280),
Senecio (123), Calceolaria (117), Baccharis (109), Palaua (108)
y Caiophora (91). Los dems tienen menos de 80 pliegos. La
especie con mayor nmero de colectas corresponde a Nolana
spathulata Ruiz & Pav., con 36 pliegos.
Representatividad geogrfca
El 96,6% (10872 pliegos) de especmenes en la coleccin
corresponden a material procedente de Per, los restantes provi-
enen de Espaa (237), Francia (107), Alemania (31), Portugal
(3) y Blgica (1). A nivel nacional, el HUSA posee colecciones
de todos los departamentos del Per con excepcin de Tumbes.
Los departamentos con mayor representacin corresponden a
Arequipa con 3375 especmenes, seguido de Pasco con 1920 y
Cusco con 996 (Fig. 1).
Figura 1. Mapa de ubicacin y nme-
ro de especmenes presentes en la
coleccin cientfca del HUSA, segn
departamento.
Figura 2. Nmero de especmenes colectados por periodos de
tiempo, en la coleccin cientfca del HUSA.
N
.
d
e
e
s
p
e
c
m
e
n
e
s
c
o
l
e
c
t
a
d
o
s
4000
3000
2000
1000
1
9
4
6
1
9
9
0
1
9
9
1
1
9
9
5
1
9
9
6
2
0
0
0
2
0
0
1
2
0
0
5
2
0
0
6
2
0
1
0
0
0
4
1 2
13
53
21
46
16
27
53
23
778
563
633
679
118
996
463
161
103
824
1920
3375
100 0 100 50
Kilmetros
DATUM: WGS84
Nmero de especmenes
segn departamento
Ubicacin de especmen
colectado
W 0 7 W 5 7 W 0 8
0
S
1
0
S
1
5
S
222
Trevio et al.
Rev. peru. biol. 19(2): 219 - 222 (Agosto 2012)
Representatividad por zonas de vida
La mayor cantidad de especmenes georeferenciados, cor-
responden a la zona de vida Bosque muy hmedo bajo tropical
con el 15,0% de estos registros, seguido por el Bosque muy
hmedo pre-montano tropical y Matorral desrtico templado
clido con el 8,1% y 6,9%, respectivamente.
Representatividad temporal
Alrededor del 90% de los especmenes de la coleccin provi-
enen de colectas realizadas a partir de 1996. Los ejemplares
peruanos ms antiguos corresponden a las especies Antochloa lep-
idula y Campyloneurum angustifolium colectados en el ao 1954.
Entre el 2001 y el 2006, el ingreso de especmenes disminuy,
sin embargo, se prioriz la implementacin del Herbario como
institucin, logrndose adems la indexacin en el ao 2004. En
el periodo 2006 - 2010 la coleccin se increment, producto del
ingreso de muestras provenientes de proyectos de investigacin y
tesis de alumnos de la Escuela de Biologa de la UNSA (Fig. 2).
Tipos de la coleccin
A continuacin se indican los tipos nomenclaturales presentes
en el Herbario HUSA y sus publicaciones ofciales:
Nolana chapiensis M.O. Dillon & Quip., Arnaldoa 14(2): 180-
184, 2007, V. Quipuscoa S. & L. Cceres M. 2495,
Dpto. Arequipa, Prov. Arequipa, Dist. Polobaya. Entre
La Gruta y el Santuario de Chapi, 2300 m, 16 45S 71
19W, 2000-11-25, Holotipo HUSA, Isotipo F.
Siphonandra nervosa Luteyn & E.M. Ortiz, J. Bot. Res. Inst.
Texas 2(1): 256, 2008, S. Vilca C., K. Arce C., C. Dva-
los M. & E. Ortiz V. 62, Puno: Carabaya, Dist. Ayapata,
near Campamento Chacayage, ca. 2600m, 13 45S 70
13W, 2004-3-10, Holotipo HUSA, Isotipo HAO, NY.
Siphonandra santa-barbarense Luteyn & E.M. Ortiz, J. Bot. Res.
Inst. Texas 2(1): 256; 2008, E. Ortiz V., S. Vilca C., C.
Arias C., S. Shua S. & H. Cristbal E. 807, Pasco:
Oxapampa, Dtto. Huancabamba, Sector Sta. Brbara,
Parque Nacional Yanachaga-Chemilln, source of the
Quebrada Cueva Blanca, 3420-3510m, 10 21 23S
75 39 20W, 2005-8-16, Holotipo HUSA, Isotipos
HOXA, MO, NY, USM.
Caiophora deserticola Weigend & Mark.Ackermann, Darwiniana
45(1): 58-61; 2007, M.Weigend & Ch.Schwarzer 7845,
Dpto. Moquegua, Prov. General Sanchez Cerro, between
Puno and Moquegua road down after junction with Lago
Desaguadero road, 3900m, 16 59S 70 42W, 2004-
4-12, Holotipo USM, Isotipos BSB, M, HUSA, HUT.
Nasa victorii Weigend, Rev. Peru. Biol. 13(1): 83. 2006,
V.Quipuscoa S., S.Leiva G. & Y.Daz 2480, Dpto. San
Martn: Prov. Huallaga, Dist. Saposoa, between El Tam-
bo and Jalca del Rayo, alt. 2500-3200 m, 2000-9-15,
Holotipo HUT, Isotipos BSB, F, HAO, HUSA.
Senecio albaniae H.Beltrn, Novon 12(1): 35. 2002, H. Beltran
3474, Lima: Huarochiri, San Pedro de Casta, Mashca,
Camino del pueblo San Pedro de Casta hacia Marcahuasi,
3680m, 11 46 S 76 35 W, 2000-5-21, Holotipo USM,
Isotipos F, HAO, HUT, MO, NY, S, TEX, US, HUSA.
Colectores y determinadores
La mayora del material de la coleccin fue colectado por
investigadores adscritos al rea de botnica y miembros del grupo
DIBIOS, principalmente Vctor Quipuscoa Silvestre (2556) y
Edgardo M. Ortiz (1268), quienes a su vez son los principales
determinadores, con 554 y 312 especmenes, respectivamente.
Agradecimientos
La informatizacin y adiciones a la coleccin no habran sido
posibles sin el fnanciamiento recibido del proyecto IABIN.
Agradecemos de igual manera a los integrantes de grupo DI-
BIOS por su participacin constante en el mantenimiento del
herbario desde sus inicios. Igualmente es muy importante desta-
car el apoyo del Dr. Abraham Calla Paredes por las facilidades
brindadas a los autores durante su gestin como Director del
herbario HUSA.
Literatura citada
Bebber D.P., M.A. Carine, J.R. Wood, et al. 2010. Herbaria are a
major frontier for species discovery. Proceedings of the
National Academy of Sciences (USA) 107: 22169 - 22171.
Cronquist A. 1981. An integrated system of classifcation of fower-
ing plants. Columbia University Press, New York.
Feeley, K.J. & M.R. Silman. 2010. Modelling the Responses of
Andean and Amazonian Plant Species to Climate Change:
The Effects of Georeferencing Errors and the Importance
of Data Filtering. Journal of Biogeography 37: 733 - 740.
Holmgren P.K. & N.H. Holmgrem. 2005. Additions to index Herb-
ariorum (Herbaria). Edition 8 - Sixteenth series. Taxon
54: 1111 - 1113.
Loiselle B.A., P.M. Jorgensen, T. Consiglio, et al. 2008. Predicting
species distributions from herbarium collections: does
climate bias in collection sampling infuence model out-
comes? Journal of Biogeography 35: 105 - 116.
Lumbsch H.T. & S.M. Huhndorf. 2007. Outline of Ascomycota.
Myconet 13: 1 - 58.
Martn-Consuegra E., J.E. Hernndez-Bermejo, M.C. Estrada, et
al. 2009. Protocolo del herbario de la Universidad de
Crdoba (COA) para la publicacin de sus datos en el
portal de GBIF: Resultados obtenidos. Boletn de la AHIM
11: 13 - 19.
Montserrat-Marti G., J.A. Ses & L. Villar. 1997. Inters de las
colecciones de herbario para los estudios de fenologa y
fenomorfologa: los ejemplos de Arbutus unedo y Phillyrea
angustifolia. Boletn de la AHIM 2: 7 - 17.
Ponder, W.F., G.A. Carter, P. Flemons & R.R. Chapman. 2001.
Anlisis Del Uso De Datos De Colecciones De Museo
En La Evaluacin De La Biodiversidad. Conservation
Biology 15: 648 - 657.
Rao R.R. 1995. Are herbaria redundant? Current Science 69: 968 - 969.
Reveal J.L. & M.W. Chase. 2011. APG III: Bibliographical Infor-
mation and Synonymy of Magnoliidae. Phytotaxa 19:
71 - 134.
Rivers, M.C., L. Taylor, N.A. Brummitt, T.R. Meagher, D.L. Roberts,
& E.N. Lughadha. 2011. How many herbarium specimens
are needed to detect threatened species? Biological Con-
servation 144: 2541 - 2547.
Schatz G.E. 2002. Taxonomy and herbaria in service of plant con-
servation: lessons from Madagascars endemic families.
Annals of the Missouri Botanical Garden 89: 145 - 152.
Squeo F.A., G. Arancio, J.R. Gutirrez, L. Letelier, M.T.K. Arroyo, P.
Len-Lobos & L. Rentera-Arrieta. 2008. Flora Amenazada
de la Regin de Atacama y Estrategias para su Conservacin.
Ediciones Universidad de La Serena, La Serena.
223
Gestin participativa de la biodiversidad con comunidades amaznicas
Rev. peru. biol. 19(2): 223 - 232 (August 2012)
Rev. peru. biol. 19(2): 223 - 232 (Agosto 2012)
Facultad de Ciencias Biolgicas UNMSM
ISSN 1561-0837
Una experiencia de gestin participativa de la biodiversidad con
comunidades amaznicas
Jos lvarez
1
y Noam Shany
2
An experience on participatory management of biodiversity with Amazonian
communities
1 Instituto de Investigaciones de la
Amazona Peruana IIAP
Av. Quiones Km. 2.5 Iquitos
Per; jalvarez@iiap.org.pe
2 Naturaleza y Cultura Internacio-
nal - NCI
COMENTARIO
Presentado: 24/01/2012
Aceptado: 12/09/2012
Publicado online: 10/11/2012
Resumen
El Instituto de Investigaciones de la Amazona Peruana IIAP, Nature and Culture International (NCI) junto con
comunidades locales, el Gobierno Regional de Loreto y otras organizaciones privadas, han estado desarrollando
en la ltima dcada un modelo de cogestin comunitaria de la biodiversidad amaznica, con resultados muy
promisorios. En el presente trabajo se analizan los principales logros de este modelo. En las tres cuencas
donde se ejecutaron estos proyectos (Tahuayo, Yanayacu del Amazonas y Nanay) observamos la reversin
de los procesos de degradacin de recursos y eco sistemas, la reduccin signifcativa de la tala, pesca y caza
ilegales, y de la cosecha destructiva de recursos; tambin han mejorado signifcativamente algunos indicado-
res econmicos de las poblaciones locales involucradas gracias a la comercializacin de productos con valor
agregado y la recuperacin de las pesqueras, la fauna silvestre y otros recursos. Las lneas maestras de
este enfoque fueron: (1) conservacin productiva (conservacin como negocio, esto es, recuperar recursos
de fora y fauna silvestres y conservarlos productivamente para la gente, generando ingresos); (2) manejo
adaptativo (medidas sencillas de manejo diseadas y adoptadas progresivamente por las propias comunida-
des con apoyo de los tcnicos, siguiendo el modelo de investigacin participativa); (3) enfoque ecosistmico
(conservacin de grandes paisajes, especialmente cuencas y ecosistemas completos, incluyendo procesos
ecolgicos priorizados, como migraciones estacionales de peces, y conservacin de reas fuente); (4) uso
y rentabilizacin del bosque en pie (priorizar generacin de ingresos de recursos silvestres manejados vs.
cambio de uso del bosque); (5) participacin de todos los actores, desde las comunidades locales organizadas
debidamente consultadas hasta las empresas privadas, las instituciones pblicas y los tomadores de decisin;
(6) desarrollo de cadenas productivas completas, con agregacin de valor a productos de la biodiversidad y
articulacin al mercado; (7) gestin adaptativa (decisiones tomadas localmente, y adecuacin progresiva de
las instituciones y normas de acuerdo a las capacidades de las comunidades).
Palabras clave: Amazonia peruana; conservacin productiva; manejo; cogestin; comunidades indgenas.
Abstract
For the past 10 years, the Instituto de Investigaciones de la Amazona Peruana (Peruvian Amazon Research
Institute) - IIAP, Nature and Culture International (NCI), together with local communities, the Regional Gover-
nment of Loreto, and other private organizations, have been developing a community co-management model
for the Amazonian biodiversity, with very promising results. In the three basins where such co-management
projects were implemented (Tahuayo, Yanayacu and Nanay), we are seeing a concrete evidence of reversals in
the processes of degradation of resources and ecosystems. In each of these basins, a signifcant reduction in
logging, illegal fshing and hunting, and destructive harvesting methods of resources can be found as a direct-
result of the co-management model. Signifcant economic improvements to local communities are achieved
mainly due to the commercialization of value-added biodiversity products, as well as the recovery of fsheries,
wildlife and other resources. The main principles of this model are : (1) productive conservation (conservation
as a business, that is to recover fora and fauna resources and their management in ways that allow income
generation); (2) adaptive management (simple management actions designed and progressively adopted by
the communities and accompanied with technical support); (3) eco-systemic approach (conservation of large
landscapes, especially watersheds, prioritized ecological processes such as fshs seasonal migrations, and
conservation of source areas); (4) making use of standing forest (to prioritize revenues from sustainable use of
forest vs. cutting down the forest); (5) participation of all stakeholders from the organized and properly consulted
local communities, to private enterprises, public institutions and all decision-makers; (6) development of complete
productive chains with value-added biodiversity products and insertion in markets; (7) adaptive management
(progressive adjustment of institutions and norms according to communities capacities).
Keywords: Peruvian Amazonia; productive conservation; management; co-management; indigenous communities.
Introduccin
Los bosques amaznicos, los ms extensos del Mundo en
los trpicos, son considerados un elemento clave en el equi-
librio climtico global, adems de una reserva estratgica de
recursos genticos y de agua dulce. Estos bosques estn mucho
ms amenazados de lo que se pensaba, y no solamente por la
deforestacin para fnes agrcolas, sino por la degradacin de
hbitat y por el cambio climtico. Nuevas tecnologas de de-
teccin remota han demostrado que la degradacin del bosque
amaznico provocada por la tala selectiva, en su mayor parte
ilegal, es mucho ms grave de lo calculado anteriormente, y
contribuye en aproximadamente un 27% a las emisiones de
carbono atribuidas a la deforestacin (Asner et al. 2010). Por
otro lado, se ha comprobado que los bosques amaznicos no
son tan resilientes como se haba pensado, y son muy suscep-
tibles a las sequas que han asolado la Amazona en los ltimos
aos (Phillips et al. 2009). La degradacin de los ecosistemas
amaznicos est provocando, a su vez, el deterioro de la calidad
de vida de las poblaciones amaznicas que dependen para su
subsistencia de los recursos de fora y fauna.
224
lvarez & Shany
Rev. peru. biol. 19(2): 223 - 232 (Agosto 2012)
Las nuevas amenazas que han surgido para los bosques
amaznicos estn relacionadas con la tala selectiva de madera,
la minera aluvial y la ampliacin de la frontera agrcola, inclu-
yendo el incremento de los cultivos de estupefacientes y, en los
ltimos aos, con los agrocombustibles. Estos ltimos, y espe-
cialmente la palma aceitera o palma africana (Elaeis guineensis)
son vistos por algunos como la mayor amenaza para los bosques
amaznicos en las prximas dcadas (junto con la soya), aunque
han sido impulsados paradjicamente para ayudar a combatir
el cambio climtico (Butler & Laurance 2009; Houtart 2011).
Especialmente preocupante es la expansin de los cultivos de
palma aceitera en la Amazona peruana: pese a que sus impulsores
plantean el uso de reas deforestadas, en la mayora de los casos
las plantaciones se establecen a costa de bosques primarios, con
frecuencia sobre territorios de uso tradicional por comunidades
locales (Smolker et al. 2007).
Pero el bosque amaznico sufre otras amenazas no tan visibles.
La defaunacin debida a la sobre caza de animales silvestres es
tan grave que puede estar poniendo en riesgo la funcionalidad
de los ecosistemas y la misma composicin de la vegetacin
(Terborgh 1999; Peres 2000; Terborgh et al. 2008). Algunos
cientfcos hablan del sndrome del bosque vaco, en alusin
a la escasez creciente de grandes animales, muchos de los cuales
cumplen un importante rol en el ecosistema como dispersores
de semillas o controladores de vegetacin (Redford 1992). La es-
casez de animales silvestres tambin tiene consecuencias sociales,
pues se est incrementando la desnutricin en las poblaciones
locales que dependen de este recurso como fuente de protena
(lvarez 2007).
Por otro lado, la creciente preocupacin por el cambio clim-
tico y la crisis econmica global ha contribuido a una aparente
disminucin de la preocupacin por la conservacin de la bio-
diversidad, a la par que ha disminuido el fnanciamiento para
proyectos orientados a esta lnea. Tambin han surgido crecientes
crticas contra un conservacionismo poco preocupado por los
temas de desarrollo y pobreza. Con frecuencia se ha impulsado
la conservacin sin tener en cuenta a los actores locales, con
nefastas consecuencias: parques de papel caros de mantener
e insostenibles en el tiempo, dinero malgastado en fracasados
proyectos conservacionistas no sostenibles. (Brown 2003; Ferraro
& Pattanayak 2006)
El aprovechamiento integral del bosque en pie (incluyendo
la produccin forestal sostenible - PFS, de productos forestales
maderables y no maderables, y de servicios ambientales, SA) es
visto como una de las alternativas para enfrentar la conversin de
bosques en reas de cultivo y pastoreo, y la degradacin por tala
selectiva y caza descontroladas. En trminos de rentabilidad, la
combinacin de estos dos usos del bosque (PFS y SA, especial-
mente el mercado de carbono) es la nica que puede competir
en rentabilidad con la conversin de bosques a cultivos de palma
aceitera (Butler & Laurance 2009).
La imparable ola de destruccin de los bosques tropicales ha
dado origen a un interesante debate en las ltimas dcadas sobre
el rol en el manejo de los recursos naturales de las comunidades
locales, especialmente las indgenas, que viven en esos bosques,
debate que ha sido llevado a las mesas de negociacin sobre el
cambio climtico; una menor atencin ha sido puesta en el rol
de las numerosas comunidades ribereas o mestizas que viven en
las riberas de los ros amaznicos ms grandes, y que por estar
ms vinculados al mercado tienen patrones de uso del territorio
algo diferentes, con ms nfasis en las actividades agropecuarias
(Anderson 1989; Goodman & Hall 1990; Davis & Wali 1994).
Se ha puesto mucho nfasis en fortalecer los derechos de los
indgenas sobre el territorio, dando por hecho que la conservacin
de los bosques se va a dar una vez que estos tengan el control total;
muchos autores que han estudiado las diversas culturas indgenas
resaltan el profundo respeto que las sociedades indgenas tradicio-
nales tienen (o ms bien, tenan) por la naturaleza y la necesidad de
preservar esas cosmovisiones y prcticas tradicionales para garanti-
zar la conservacin de los bosques, ignorando en cierto modo los
incontenibles y cada vez ms acelerados procesos de aculturacin e
insercin en la economa de mercado (Redford & Stearman 1993).
Sin embargo, la gran mayora de los estudios sobre comunidades
indgenas ha sido hecha en comunidades muy tradicionales, y se ha
dejado de lado las comunidades ms vinculadas al mercado y, por
tanto, ms aculturadas y menos conservacionistas, aun cuando
el proceso de trnsito de unas a otras es cada vez ms acelerado, y
aparentemente irreversible e inevitable (Johnson 1989; Holt 2005;
De Soto 2010). La experiencia demuestra que tambin los indge-
nas pueden ser partcipes de destruccin de los bosques tropicales,
o al menos ser testigos pasivos, cuando son dejados a merced de
las fuerzas de la globalizacin sin instrumentos tcnicos y legales
para gestionar sus bosques y benefciarse econmicamente de ellos
(Redford & Taber 2000).
Algunos autores ya hace tiempo hicieron notar que los ind-
genas pueden ser aliados de la conservacin siempre y cuando,
adems de control efectivo y la seguridad legal de sus territo-
rios, dispongan de capacitacin y herramientas tcnicas para el
manejo sostenible y rentable de sus recursos (Clad 1984; Davis
& Wali 1994; Peres 1994). Un reciente estudio demuestra que
los bosques en manos de comunidades locales (en 16 pases de
Amrica Latina, frica y Asia) sufren tasas de deforestacin sig-
nifcativamente menores que aquellos dentro de reas naturales
protegidas de forma estricta: las reas protegidas perdieron en
promedio 1,47% de su cobertura boscosa al ao, en compa-
racin con 0,24% de los bosques manejados por comunidades
(Porter-Bolland et al. 2011).
Debemos partir del hecho de que las culturas y sociedades
indgenas y mestizas amaznicas no son estticas, como no lo son
sus usos de los recursos silvestres y sus formas de organizacin.
La variabilidad ecolgica y la dinmica histrica han impulsado
a las sociedades indgenas a adaptarse y a crear constantemente
formas nuevas de uso del territorio y los recursos (Clay 1988;
Posey & Balee 1989). Sus sistemas de creencias, que regulaban
en el pasado el uso del territorio y los recursos, a travs de tabes
y otros mecanismos, se enfrentan con desventaja al avasallador
avance de la cultura y economa occidentales, y terminan colap-
sando si las comunidades indgenas no encuentran alternativas
econmicas social, ambiental y econmicamente sostenibles.
Muchos proyectos de conservacin en la Amazona han fraca-
sado justamente en el mediano plazo, o bien por desconocer
las particularidades socioculturales de las comunidades involu-
cradas, o por descuidar el aspecto de sostenibilidad econmica,
generalmente en temas de valor agregado y mercado (Gasch
2004; lvarez 2007).
La creciente emigracin de las zonas rurales a la ciudades
grandes y medianas ha sido vista como una esperanza para la
conservacin de los bosques tropicales (Wright & Muller-Landau
225
Gestin participativa de la biodiversidad con comunidades amaznicas
Rev. peru. biol. 19(2): 223 - 232 (August 2012)
2006); sin embargo, el acelerado crecimiento de las ciudades
amaznicas est generando grandes impactos en las reas ms
cercanas, y no est mitigando la sobre explotacin de los recur-
sos de fora y fauna en las zonas alejadas, adonde extractores no
residentes siguen con sus actividades para abastecer la demanda
de las ciudades, generalmente sin manejo y con prcticas des-
tructivas de cosecha (Parry et al. 2010).
Comunidades locales, aliadas de la conservacin
Ante las altas tasas de deforestacin y la imbatible disminu-
cin y fragmentacin de los bosques tropicales prstinos, algunos
dan por hecho que no podrn ser conservadas grandes exten-
siones de bosques primarios, y ponen nfasis en la conservacin
de los agroecosistemas, especialmente los mosaicos de hbitat
relacionados con la agricultura tradicional (Vandermeer &
Perfecto 2007b). Quienes se han resignado a perder las ltimas
grandes extensiones de los bosques tropicales estn abogando
por nuevas e innovadoras formas de conservar en un escenario
dominado por comunidades y pequeos agricultores con ttulos
de propiedad sobre bosques y reas agrcolas: plantean conservar
no solo fragmentos de los bosques remanentes (incluyendo las
reas protegidas), sino la matriz que incluye reas manejadas
de bosques y reas de cultivo tradicionales favorables para la
biodiversidad (sistemas agroforestales y otros; Roe & Jack 2001;
Schwartzman & Zimmerman 2005; Vandermeer & Perfecto
2007a). Se impone la idea de que las alianzas y colaboracin con
las comunidades locales y con los movimientos y organizaciones
de campesinos e indgenas ser crucial para la conservacin de la
biodiversidad en general y de los bosques tropicales en particular
(Vandermeer & Perfecto 2007b). Aunque los objetivos de estos
movimientos suelen estar vinculados principalmente a la defensa
de sus derechos sobre la tierra y otros recursos (agua, bosques,
etc.), el tema ecolgico est cada vez ms presente, y en sus
mensajes y acciones aparece la conservacin de los ecosistemas
y la biodiversidad como un objetivo estrechamente ligado con la
defensa de su modo de vida (ver por ej. Wright & Wolford 2003).
Cabe destacar que en la Regin Loreto el escenario es fa-
vorable todava para la conservacin y manejo sostenible de
grandes extensiones de bosques primarios: apenas un 3% de las
36.8 millones de hectreas de bosques han sido transformados
por la agricultura.
Por otro lado, existe una visin bastante pesimista de que la
conservacin de los bosques tropicales no podr sostenerse en
argumentos econmicos (el bosque amaznico vale ms vivo
que muerto) y las opciones de desarrollo siempre son priorizadas
frente a las de conservacin, frente a lo que se plantea el fortale-
cimiento de los argumentos ticos (Terborgh 1999; Sanderson
& Redford 2003).
En un escenario en que disminuyen los fondos para con-
servacin a la par que se incrementan las amenazas contra los
bosques amaznicos, las comunidades locales son vistas cada vez
ms no tanto como una amenaza sino como potenciales aliados
de la conservacin, como parte de la solucin en vez de parte del
problema (Vandermeer & Perfecto 2007a; Vermeulen & Sheil
2007). En los ltimos aos se ha puesto de moda en enfoque de
la participacin en los proyectos y actividades de conservacin
en pases tropicales; la mayora de ellos, sin embargo, y a pesar
de la retrica participativa, siguen siendo diseados y contro-
lados por organizaciones y pequeos grupos de expertos; en ellos
siguen estando ausentes tanto la consulta formal como las formas
democrticas en la ejecucin de los proyectos, al tiempo que se
ignoran en buena medida las iniciativas, prioridades e intereses
de las comunidades locales (Campbell & Vainio-Mattilia 2003;
Vermeulen & Sheil 2007).
La participacin se ha convertido en algunos casos en una
especie de ritual que todo proyecto incluye sin mayor criterio, y
puede convertirse en un elemento distorsionador: la participacin
de las comunidades tanto en el cuidado de los recursos como en
el monitoreo no es sostenible si los pobladores locales no ven
retribuido su esfuerzo con benefcios claros y tangibles (Cooke &
Kothari 2001). Si bien muchos conservacionistas estn de acuerdo
en la necesidad de establecer alianzas con las comunidades locales
en la bsqueda de objetivos comunes y lneas coincidentes para
conservar la biodiversidad y los ecosistemas tropicales, hay mucha
polmica sobre el cmo. Los joint ventures y otros negocios
compartidos con comunidades han sido vistos como una de
las mayores oportunidades para la conservacin y el desarrollo
sostenible en bosques tropicales (CSD 2004).
Un intenso debate tambin ha surgido en torno a la necesidad
de establecer alianzas (partnerships) entre comunidades ind-
genas, cientfcos y profesionales de la conservacin, gobiernos
y agencias de cooperacin internacional (Vermeulen & Sheil
2007). Los bionegocios con base en los incalculables recursos
genticos amaznicos y el incipiente mercado de carbono se
avizoran como muy positivas oportunidades para impulsar
productivas y estratgicas alianzas entre las comunidades locales,
empresas, gobierno y organizaciones conservacionistas y de co-
operacin, en la bsqueda de un desarrollo realmente inclusivo y
sostenible. Sin embargo, estas alianzas deben que tener objetivos
claros compartidos, altos estndares de participacin incluyen-
do decisiones, derechos y deberes, y riesgos compartidos- y deben
benefciar claramente a todas las partes, y especialmente a las
comunidades locales involucradas (Campbell & Vainio-Mattilia
2003; Vermeulen & Sheil 2007).
Existen diversas y promisorias experiencias de colaboracin
entre cientfcos conservacionistas y comunidades, por ejemplo
en el tema de manejo de fauna silvestre y en monitoreo de la
biodiversidad (Bodmer et al. 1997). Pese a que algunos cient-
fcos ponen en cuestin su precisin cientfca, el monitoreo
comunitario o participativo con comunidades ha demostrado
ser sumamente efectivo en trminos de costo-resultados y de
impacto en la gestin, que en defnitiva es lo que ms importa.
Adems, las comunidades involucradas en monitoreo suelen
mejorar el control sobre sus territorios y recursos, lo que con-
tribuye a la conservacin de la biodiversidad (Getz et al. 1999;
Sheil & Lawrence 2004; Danielsen et al. 2007).
Las estrategias de conservacin de los ecosistemas a travs de la
creacin de reas protegidas (especialmente parques nacionales)
y compra de tierras estn llegando a su techo y se manifestan
insostenibles, debido a los excesivos costos sociales y econmicos
(Ghimire & Pimbert 1997; Bruner et al., 2001; Balmford &
Whitten 2003). Muchas de las iniciativas de conservacin se
ven frenadas por la oposicin de comunidades locales y orga-
nizaciones que las apoyan, porque no han sido consultadas de
forma adecuada, y porque se sienten limitadas en sus aspiraciones
de acceso a los recursos silvestres; tambin se oponen sectores
econmicos interesados en el aprovechamiento de recursos, que
ven en las reas protegidas una amenaza a sus intereses.
226
lvarez & Shany
Rev. peru. biol. 19(2): 223 - 232 (Agosto 2012)
En la Amazona peruana en particular, la cobertura de las
formas tradicionales de reas protegidas est llegando a su lmite
(aproximadamente un 20% del territorio, y en algunas regiones,
como Madre de Dios, an ms) y debe buscarse alternativas para
proteger grandes extensiones de bosques todava en el limbo legal
de reas de libre disponibilidad del Estado, donde persiste bue-
na parte de la biodiversidad. Adems, cada vez es ms evidente
que la conservacin de la biodiversidad amaznica y los servicios
y recursos que proveen los bosques amaznicos requieren del
mantenimiento de grandes y continuas extensiones de bosques
y la proteccin de los procesos ecolgicos y evolutivos que hacen
posible la biodiversidad (Garca Villacorta 2010).
En la Regin Loreto, en la Amazona norperuana, tenemos
una oportunidad histrica para conservar grandes extensiones
de bosques tropicales porque aqu todava los bosques valen ms
vivos que muertos y se conserva todava el 97% de los bosques
originales, 36.8 millones de hectreas en total. La frontera agrco-
la es incipiente, y gran parte de las cerca de 3,000 comunidades
locales, tanto indgenas como ribereas, basan su economa en el
aprovechamiento sostenible de los recursos forestales y de fauna
silvestre (Pyhl 2003; lvarez 2006).
En este documento se describen algunas experiencias que
demuestran que el bosque amaznico puede valer ms vivo que
muerto, en pie que talado, contradiciendo la pesimista visin
de algunos expertos, incluyendo la de J. Terborgh (1999). In-
novadoras formas de concebir las reas protegidas como zonas
de aprovechamiento sostenible de recursos por comunidades
locales, manteniendo la integridad y salud de los ecosistemas,
han cambiado en cierto modo el escenario, abriendo la posibi-
lidad de crear nuevas reas en alianza con comunidades locales
interesadas en conservar las zonas fuente de sus recursos de fauna
y fora (Bodmer 2000; Bodmer & Puertas 2000).
Una visin tergiversada de la Amazona
En las ltimas dcadas se han invertido decenas de millones
de dlares de la cooperacin internacional y del presupuesto
pblico del Per para impulsar proyectos de desarrollo y con-
servacin con comunidades locales amaznicas, con resultados
irrisorios, o en muchos casos contraproducentes. Las amenazas
contra biodiversidad crecen da a da, a la par que se deteriora la
calidad de vida de las poblaciones amaznicas que dependen de
los recursos de fora y fauna para su subsistencia. Los ndices de
pobreza y desnutricin en la selva son los peores del Per, y en
algunos casos han empeorado, a contracorriente de lo que ocurre
en la mayor parte del pas, donde ha ocurrido una signifcativa
reduccin de la pobreza.
El fracaso de la mayora de los proyectos (tanto de conserva-
cin como de desarrollo) se debe -a nuestro juicio y de algunos
autores- a la falta de comprensin tanto de la realidad ecolgica
de la Amazona baja como de las sociedades amaznicas que viven
en ella (Gasch 2004). Parte del problema reside en el divorcio
entre quienes impulsan la conservacin de la biodiversidad y los
que impulsan el desarrollo de las comunidades (Guha 1997; Holt
2005). Por ejemplo, la mayora de los proyectos de desarrollo
con comunidades impulsaron actividades agropecuarias copiadas
de otras realidades, no apropiadas para los pobres y complejos
suelos amaznicos, ni acordes con la tradicin extractivista y
las formas de organizacin de los pobladores locales (CADMA
1992; lvarez 2007).
Para citar un ejemplo: desde un tiempo a esta parte se ha dado
en promover en Amazona proyectos productivos comunales,
cuando la comunidad no es una unidad social de produccin,
y el modo de produccin tradicional es familiar, o de familias
extensas (Gasch 1999; 2010). De este modo han fracasado cien-
tos de proyectos de piscigranjas familiares, crianzas comunales
de animales domsticos, chacras comunales, etc.
La Amazona peruana ha sido histricamente abordada desde
el Gobierno Nacional asentado en la costa (Lima, la capital) con
una visin sesgada de su realidad ecolgica y social. No solo se
trata de los mitos sobre la Amazona que an sobreviven en el
imaginario colectivo nacional (mitos de la uniformidad, del vaco
amaznico, de la fertilidad de los suelos CADMA 1992) sino
de una visin del desarrollo calcada de otras realidades, ajenas a
los ecosistemas y las sociedades amaznicas. Pero la Amazona es
diferente: primero, por su extraordinaria riqueza natural y cul-
tural, que se expresa en su extremada diversidad de ecosistemas,
especies genes y culturas humanas; y segundo, por su extremada
fragilidad. El manejo de la biodiversidad en Amazona enfrenta
retos y problemas diferentes a los de otras latitudes. Por ejemplo,
son escasas y poco efectivas las medidas y tcnicas de manejo
aplicables en los complejos ecosistemas amaznicos (y aplicadas
con xito en pases templados), debido especialmente al rgimen
de libre acceso a los recursos, a la inaccesibilidad de muchas reas
de extraccin, a la enorme variabilidad ecolgica de los ecosistemas
amaznicos, al escaso conocimiento disponible sobre historia natu-
ral, densidad y dinmica poblacional de la mayora de las especies,
a la idiosincrasia de las poblaciones locales, y a las limitaciones de
las instituciones responsables de hacer cumplir la ley.
Los funcionarios pblicos y miembros de organizaciones de
cooperacin que se han acercado a la Amazona tratando de
impulsar proyectos de desarrollo la han visto generalmente como
un suelo a cultivar, y no han podido ver el bosque (o han visto
solamente madera comercial en l), ni las formas tradicionales de
aprovecharlo que tienen las comunidades locales. Desde tiempos
del Presidente Belande Terry (en sus dos gobiernos, a fnes de
los 60 y principios de los 80), con su lema Tierra sin gente para
gente sin tierra, y su visin de la Amazona como arroz con
bisteck, se ha impulsado proyectos agropecuarios calcados de
la sierra o la costa del Per, absolutamente inapropiados para
los suelos pobres y cidos de la selva baja, y no acordes con la
cultura y formas de produccin de las comunidades amaznicas
tradicionales (CIAM 2011).
No es por eso extrao que la Amazona peruana haya sido
acertadamente califcada como un cementerio de proyectos. No
hay comunidad indgena o riberea que no tenga en su historia
algn proyecto de desarrollo fracasado, sea impulsado por alguna
agencia del Gobierno o por alguna organizacin no gubernamen-
tal. La mayora de estos proyectos han estado y estn relacionados
con la produccin agrcola o pecuaria, incluyendo promocin
de cultivos comerciales, reforestacin con especies nativas o
introducidas, crianza de animales (menores o mayores), tanto
nativos como introducidos, piscicultura familiar o comunal, y
otros similares. En otros casos son las obras de infraestructura,
con bastante frecuencia sin un sentido productivo ni el mnimo
sentido de la prioridad: plazas, veredas, losas deportivas, puentes
sobre minsculas quebradas
El enfoque excesivamente tcnico de la mayora de los pro-
yectos, ajeno en buena medida a las aspiraciones, capacidades
227
Gestin participativa de la biodiversidad con comunidades amaznicas
Rev. peru. biol. 19(2): 223 - 232 (August 2012)
y visin de la poblacin local, ha sido identifcado como otra
de las causas ms comunes de fracasos de proyectos, los que
suelen ser dirigidos por profesionales muy especializados con
una visin muy sesgada de la realidad (Gasch 2004; Santoyo
2004) . Sin embargo, tambin ciertas visiones antropolgicas,
con su excesivo -y con frecuencia exclusivo- nfasis en aspectos
socioculturales, y con limitado conocimiento e inters en temas
ecolgicos y productivos, han ayudado poco a promover modelos
de desarrollo sostenible en las comunidades amaznicas.
Por otro lado, la mayora de los proyectos de desarrollo y
conservacin han sido concebidos con un muy limitado hori-
zonte espacial y temporal: se trata de intervenciones en una sola
o un puado de comunidades, con frecuencia no relacionadas
espacialmente, y de corto plazo; en los aspectos productivos, la
mayora no consideran la cadena productiva completa comen-
zando del aprovechamiento sostenible del recurso-, y menos an
aspectos de gestin empresarial y de mercado. Esto determina
casi de antemano su insostenibilidad. Para garantizar la sosteni-
bilidad ecolgica es necesario gestionar con enfoque ecosistmico
grandes espacios, preferiblemente cuencas completas, y para
garantizar sostenibilidad econmica debe promoverse la agre-
gacin de valor, el fortalecimiento de capacidades empresariales
y la articulacin con los mercados.
Por poner algunos ejemplos del tema ecolgico: no tiene
mucho sentido, ni impacto ciertamente, tratar de manejar so-
lamente un pequeo lago amaznico, y menos una sola especie
comercial del lago, cuando la mayora de los peces son grandes
emigrantes, y tambin necesitan el bosque inundable circun-
dante y de otros ecosistemas fuera del lago para alimentarse y
reproducirse. Muchas especies de fauna requieren de grandes
territorios para mantener poblaciones saludables y viables: no
es posible manejar, por ejemplo, ungulados o grandes primates
en el territorio de una sola comunidad, por muy grande que sea,
porque no alberga poblaciones viables; es indispensable, por eso,
trabajar con una visin de paisaje, integrando grandes espacios
y trabajando con todas o las mayora de las comunidades y sus
organizaciones representativas de una cuenca, as como con otros
actores que intervienen en una cuenca o zona determinada. Hoy
se sabe que para que la caza de animales silvestres amaznicos
sea sostenible es necesario proteger zonas fuente conectadas
con las zonas de caza, donde los animales se puedan reproducir
y dispersarse hacia las zonas de caza (Bodmer 2000; Ferraro &
Pattanayak 2006)
La falta de sostenibilidad ambiental tiene otras caras: la
mayora de los proyectos de desarrollo en Amazona peruana
centran sus esfuerzos en promover alternativas productivas con
base en especies cultivadas o criadas en cautividad (cultivos na-
tivos o, con frecuencia, exticos, y crianza de animales exticos,
incluyendo ganadera extensiva), promoviendo con ello directa o
indirectamente la desvalorizacin del bosque y la deforestacin.
Son muy escasos los proyectos e iniciativas, tanto de las insti-
tuciones del Estado como de la cooperacin internacional, que
busquen generar recursos econmicos aprovechando el bosque
en pie (esto es, manejo de recursos silvestres de fora y fauna). Sin
embargo, la mayor parte de los pobladores rurales amaznicos
siguen usando en gran medida los recursos silvestres, forestales
y acuticos, para fnes de subsistencia y tambin para obtener
recursos econmicos con su venta; por ejemplo, el pescado y la
carne de animales silvestres representan entre el 70 y el 80% de
la ingesta de protena en buena parte de las familias rurales en
Loreto. Un estudio realizado en comunidades del bajo Nanay,
bien conectadas con el mercado de Iquitos, mostr que alrede-
dor del 70% de los ingresos familiares provienen de la venta de
recursos silvestres, especialmente recursos forestales maderables
y no maderables, carne de monte, frutos silvestres y pescado
(Pyhl 2003). Solamente en las comunidades muy cercanas a
las ciudades y pueblos grandes, y en las riberas de los grandes
ros de agua blanca (con suelos aluviales relativamente frtiles) la
agricultura representa un porcentaje importante de los ingresos.
Trabajando con comunidades locales y conser-
vando los bosques
Desde fnales de la dcada de 1990, el Instituto de Investigacio-
nes de la Amazona Peruana IIAP, consciente de esta situacin y
ante la imparable degradacin de los recursos naturales renovables
y de la calidad de vida de las poblaciones amaznicas, se propuso
impulsar proyectos que buscasen combinar la conservacin de la
diversidad biolgica y de los ecosistemas y el desarrollo sostenible
de la poblacin, con una slida base ecolgica y socioeconmica,
con visin ecosistmica y de grandes paisajes, y con participacin
activa de las comunidades locales involucradas.
El primer proyecto impulsado por el IIAP se ejecut entre el
ao 2000 y el 2004 con apoyo del Banco Mundial (y con fondos
del Global Environmental Fund) en la cuenca del ro Nanay,
Loreto, Per. El IIAP se plante el reto de experimentar un
modelo de desarrollo sostenible y conservacin de la diversidad
biolgica en una cuenca emblemtica de la Amazona peruana;
se escogi el Nanay por ser ejemplo de la extremada riqueza
biolgica y variabilidad de ecosistemas de la regin, y por pre-
sentar un panorama extremo de depredacin y degradacin de
los ecosistemas, dada su cercana y conectividad con Iquitos. Se
saba de muchos proyectos fallidos de desarrollo en esta cuenca.
La experiencia del Proyecto Conservacin de la biodiversidad
y manejo comunal de los recursos naturales en la cuenca del ro
Nanay (Proyecto Nanay) demuestra que s es posible aplicar con
xito medidas de conservacin de la diversidad biolgica y de
desarrollo sostenible con comunidades locales, con las siguientes
estrategias de intervencin: 1) Enfoque integral y de cuenca:
manejo integral del ecosistema, preferible al nivel de cuenca; 2)
enfoque de conservacin productiva: conservar para la gente,
generar riqueza sin alterar los ecosistemas; la poblacin conserva
lo que le es til; 3) defnicin clara de derechos de acceso a los
recursos y, especialmente, control del territorio por parte de
comunidades; 4) diagnstico participativo de la problemtica
de los recursos naturales; 5) apoyo a la organizacin interna
de las comunidades, capacitadas y fortalecidas para el manejo
(asambleas y grupos comunales de manejo), y a la organizacin
supracomunitaria (comits zonales de gestin, federaciones); 6)
diseo participativo y aplicacin de planes de manejo adaptativo
al nivel de comunidad; 7) co-gestin, aplicacin de reglamentos
comunales internos de acceso a los recursos con apoyo de los
tcnicos y las instituciones del Estado; 8) formalizacin / legaliza-
cin del aprovechamiento de los recursos; 9) alianzas estratgicas
con instituciones pblicas y organizaciones de cooperacin; y
10) diversifcacin de la base productiva y acceso al mercado
(IIAP 2004; lvarez 2006).
En la Tabla 1 se muestra un ejemplo de los avances conse-
guidos en la disminucin de amenazas en 16 comunidades de
228
lvarez & Shany
Rev. peru. biol. 19(2): 223 - 232 (Agosto 2012)
la cuenca, en este caso la comunidad nativa San Juan de Ungu-
rahual. Estos datos fueron recogidos por las mismas comunidades
a travs del empleo de matrices de monitoreo comunitario.
Entre el 2004 y el 2007 el IIAP ejecut otro proyecto en la
parte baja de la misma cuenca, incluyendo la Reserva Nacional
Allpahuayo-Mishana, esta vez fnanciado por el Gobierno de
Finlandia con fondos de canje de deuda por naturaleza: Proyecto
Diversidad Biolgica de la Amazona Peruana Per Finlandia
(BIODAMAZ Fase II; lvarez et al. 2004). Las lneas maestras
de trabajo con comunidades locales replicaron y mejoraron las
lneas estratgicas aplicadas en el Proyecto Nanay, especialmente
el enfoque de co-gestin, dado que este proyecto se ejecut en
el escenario de un rea protegida, donde las comunidades com-
parten las competencias de gestin de recursos naturales con los
tcnicos y representantes del Estado. Durante esta experiencia
tambin se logr grandes avances en la aplicacin del modelo de
manejo comunal de recursos de la biodiversidad, con resultados
notables en la disminucin de las amenazas para la biodiversi-
dad y la degradacin de los ecosistemas. En las Figuras 1 y 2 se
aprecia, por ejemplo, la signifcativa disminucin del ingreso
de extractores ilegales de los recursos ms importantes para la
economa local (hojas de la palmera irapay y de la madera re-
donda de varillales) en la Reserva Nacional Allpahuayo-Mishana
(tambin esta informacin es fruto de la aplicacin de matrices
de monitoreo comunitario).
Recurso/actividad N de extractores ilegales de:
Ao 2002 2003 2004 Disminucin (%)
Irapay 164 133 24 85
Madera redonda 201 249 44 82
Madera aserro 185 162 20 89
Pesca 514 841 224 73
rboles maderables para lea 72 63 16 77
Cosecha destructiva de RR.NN. (%)
Ao 2002 2003 2004
Ungurahui 100 50 10 90
Aguaje 100 49 9 91
Irapay 74 31 7 51,3
Madera redonda 54 32 6.3 70,3
Pesca destructiva 29 16 0 55,1
Tala ilegal 36 21 3 66,6
Tabla 1. Disminucin de las amenazas para los recursos naturales en 16 comunidades de la cuenca del Nanay.
0
4
8
12
16
20
A
n
g
u
i
l
l
a
M
i
s
h
a
n
a
P
o
r
v
e
n
i
r
S
a
n
M
a
r
t
n
Y
u
t
o
A
n
g
u
i
l
l
a
M
i
s
h
a
n
a
P
o
r
v
e
n
i
r
S
a
n
M
a
r
t
n
Y
u
t
o
A
n
g
u
i
l
l
a
M
i
s
h
a
n
a
P
o
r
v
e
n
i
r
S
.
M
Y
u
t
o
A
n
g
u
i
l
l
a
M
i
s
h
a
n
a
P
o
r
v
e
n
i
r
S
.
M
Y
u
t
o
2003 2004 2005 2006
N
m
e
r
o
d
e
i
n
f
r
a
c
t
o
r
e
s
Comunidades -aos
Figura 1. ndice de extractores ilegales de irapay en comunidades de
la Reserva Nacional Allpahuayo-Mishana (RNAM) (Fuente: Informes
fnales del Proyecto BIODAMAZ Fase II. IIAP, Iquitos, 2007. http://
www.iiap.org.pe/biodamaz/faseii/avances/Doc_tec.htm)
0
5
10
15
20
25
30
35
A
n
g
u
i
l
l
a
M
i
s
h
a
n
a
P
o
r
v
e
n
i
r
S
a
n
M
a
r
t
n
Y
u
t
o
A
n
g
u
i
l
l
a
M
i
s
h
a
n
a
P
o
r
v
e
n
i
r
S
a
n
M
a
r
t
n
Y
u
t
o
A
n
g
u
i
l
l
a
M
i
s
h
a
n
a
P
o
r
v
e
n
i
r
S
.
M
Y
u
t
o
A
n
g
u
i
l
l
a
M
i
s
h
a
n
a
P
o
r
v
e
n
i
r
S
.
M
Y
u
t
o
2003 2004 2005 2006
N
m
e
r
o
d
e
i
n
f
r
a
c
t
o
r
e
s
Comunidades-aos
Figura 2. ndice de infractores de madera redonda en comunidades
de la la Reserva Nacional Allpahuayo-Mishana (RNAM) (Fuente:
Informes fnales del Proyecto BIODAMAZ Fase II. IIAP, Iquitos, 2007.
http://www.iiap.org.pe/biodamaz/faseii/avances/Doc_tec.htm)
0
50 50
30
40
20
0
22
15
0
40
0 0
6
0 0 0
20
0 0
9
0 0
0
10
20
30
40
50
60
A
n
g
u
i
l
l
a
M
i
s
h
a
n
a
P
o
r
v
e
n
i
r
S
.
M
a
r
t
n
Y
u
t
o
1
5
d
e
A
b
r
i
l
A
n
g
u
i
l
l
a
M
i
s
h
a
n
a
P
o
r
v
e
n
i
r
S
.
M
a
r
t
n
Y
u
t
o
1
5
d
e
A
b
r
i
l
A
n
g
u
i
l
l
a
M
i
s
h
a
n
a
P
o
r
v
e
n
i
r
S
.
M
a
r
t
n
Y
u
t
o
1
5
d
e
A
b
r
i
l
A
n
g
u
i
l
l
a
M
i
s
h
a
n
a
P
o
r
v
e
n
i
r
S
.
M
a
r
t
n
Y
u
t
o
2003 2004 2005 2006
P
o
r
c
e
n
t
a
j
e
Aos y comunidades
Figura 3. Porcentaje de familias que efectan prcticas destructivas
de cosecha de irapay.
En esta reserva se produjo tambin una substancial disminu-
cin de las prcticas de cosecha destructiva entre los pobladores,
otra de las amenazas ms importantes para la comunidad y para la
biodiversidad. Como muestra presentamos el caso de la palmera
irapay y madera redonda, los dos recursos ms importantes en
trminos econmicos para las comunidades de la zona (Figs. 3
y 4; lvarez et al. 2004).
Como resultado de la aplicacin de medidas de manejo adap-
tativo por las comunidades locales se produjo una substancial
229
Gestin participativa de la biodiversidad con comunidades amaznicas
Rev. peru. biol. 19(2): 223 - 232 (August 2012)
recuperacin del recurso pesquero en los cuerpos de agua de la
Reserva Nacional Allpahuayo Mishana - RNAM, tal como se
puede comprobar en la Figura 5, que mide el promedio de peces
capturados por faena con distintas tcnicas de pesca (captura
por unidad de esfuerzo). Las medidas de manejo, propuestas
por las mismas comunidades y aprobadas en acta de asamblea
comunal incluyeron, entre otros aspectos: prohibicin de pesca
comercial por forneos, control de mtodos destructivos de pesca
(txicos y explosivos, redes no selectivas, honderas y arrastrado-
ras), regulacin del tamao de la malla de redes-trampa, lmites
en el volumen de capturas por pescador con fnes comerciales,
prohibicin de pesca comercial durante la poca de desove, y
control de tala de bosques ribereos (lvarez et al. 2004).
Cabe destacar que solo se monitore directamente las capturas
pesqueras porque este recurso se recupera de forma relativamente
rpida, a diferencia de la fauna silvestre terrestre y de la fora, y
es factible monitorear las capturas por unidad de esfuerzo traba-
jando con los pescadores locales. En relacin con otros recursos,
se opt por monitorear solo las intervenciones de la poblacin
sobre el recurso, y no directamente el recurso, dado que: i) el
monitoreo directo es sumamente caro y de mediano/largo plazo,
y fuera del alcance por tanto del horizonte de un tpico proyecto
de cooperacin al desarrollo y la conservacin; y ii) los cambios
en la composicin y abundancia de los recursos son difcilmente
distinguibles en el corto plazo de las fuctuaciones poblacionales
naturales (de la fauna silvestre en especial) o del ruido estadstico,
y por tanto son irrelevantes para los objetivos de un proyecto. El
enfoque del monitoreo ha sido netamente participativo: las matri-
ces de monitoreo fueron completadas con participacin directa de
las mismas comunidades, unas veces en asamblea comunal, otras
en reuniones con lderes destacados de la comunidad. Si bien la
informacin refejada en las matrices de monitoreo comunitario
-y en las fguras aqu presentadas- es de grano grueso, porque se
basa en datos referidos por la poblacin, es muy til y relevante
para detectar tendencias en los patrones de uso de recursos de las
comunidades, y para monitorear la disminucin de las amenazas.
No nos cabe ninguna duda de que son un indicador fel del estado
de conservacin real de los recursos de fora y fauna silvestre.
Finalmente, entre el 2007 y el 2010 se ha estado ejecutando
un proyecto de apoyo a la creacin y gestin de reas de con-
servacin regional en la Regin Loreto, con fnanciamiento de
Fundacin Moore (Proyecto de Apoyo al PROCREL, Programa
de Conservacin, Gestin y Uso Sostenible de la Biodiversidad
en la Regin Loreto). El Proyecto fue co-ejecutado por el IIAP,
el Gobierno Regional de Loreto, y la organizacin no guberna-
mental Nature and Culture International - NCI. La primera rea
de conservacin regional en ser creada y gestionada fue el rea
de Conservacin Regional Comunal Tamshiyacu-Tahuayo, en el
ao 2007, aunque se trabaj con las comunidades desde varios
aos antes. Al ao siguiente fueron creadas otras dos, el ACR
Ampiyacu-Apayacu, y el ACR Alto Nanay-Pintuyacu-Chambira.
Los logros de la aplicacin del modelo de co-gestin comunal
han sido muy signifcativos, especialmente en el ACR Tamshiyacu-
Tahuayo, donde las comunidades ya llevaban aos gestionando
esta zona, an antes de ser declarada ofcialmente como rea
de conservacin regional. Con apoyo de organizaciones como
Wildlife Conservation Society las comunidades de la cuenca
del ro Tahuayo han estado aplicando por ms de una dcada
medidas de manejo de fauna silvestre con gran xito (Bodmer
et al. 1997; Bodmer & Puertas 2000, Novaro et al. 2000), y el
modelo ha sido reconocido como uno de los mejores en gestin
de bosques tropicales a nivel mundial (Congreso Forestal Mundial
2009, Buenos Aires). En el Per tambin han recibido diversos
reconocimientos y premios. El Proyecto de Apoyo al PROCREL
impuls el manejo comunal de otros recursos adems de la fauna
silvestre, incluyendo pesqueras, palmera irapay (Lepidocaryum
tenue), palmera chambira (Astrocaryum chambira), palmera aguaje
(Mauritia fexuosa), camu camu (Myrciaria dubia), entre otros
recursos importantes para la poblacin.
Las comunidades no solo han conseguido recuperar la
productividad de sus bosques y ecosistemas acuticos, y han
mejorado su calidad de vida gracias a la abundancia de fauna
silvestre, pescado y otros recursos: tambin han mejorado subs-
0
60
40 40 40
50
0
15 15
0
40
5
0
6
0 0 0
20
0 0 0
13
0
0
10
20
30
40
50
60
70
A
n
g
u
i
l
l
a
M
i
s
h
a
n
a
P
o
r
v
e
n
i
r
S
.
M
a
r
t
n
Y
u
t
o
1
5
d
e
A
b
r
i
l
A
n
g
u
i
l
l
a
M
i
s
h
a
n
a
P
o
r
v
e
n
i
r
S
.
M
a
r
t
n
Y
u
t
o
1
5
d
e
A
b
r
i
l
A
n
g
u
i
l
l
a
M
i
s
h
a
n
a
P
o
r
v
e
n
i
r
S
.
M
a
r
t
n
Y
u
t
o
1
5
d
e
A
b
r
i
l
A
n
g
u
i
l
l
a
M
i
s
h
a
n
a
P
o
r
v
e
n
i
r
S
.
M
a
r
t
n
Y
u
t
o
2003 2004 2005 2006
P
o
r
c
e
n
t
a
j
e
Aos y comunidades
Figura 4. Porcentaje de comuneros que efectan malas prcticas para el recurso madera redonda (Fuente: Informes fnales del Proyecto
BIODAMAZ Fase II. IIAP, Iquitos, 2007. http://www.iiap.org.pe/biodamaz/faseii/avances/Doc_tec.htm)
13
14
20
24
31
0
5
10
15
20
25
30
35
2002 2003 2004 2005 2006
N
p
r
o
m
e
d
i
o
Aos
Figura 5. Evolucin del nmero promedio de ejemplares pescados en
la RNAM empleando diferentes tcnicas de pesca (Fuente: Informes
fnales del Proyecto BIODAMAZ Fase II. IIAP, Iquitos, 2007. http://
www.iiap.org.pe/biodamaz/faseii/avances/Doc_tec.htm)
230
lvarez & Shany
Rev. peru. biol. 19(2): 223 - 232 (Agosto 2012)
tancialmente su economa, gracias al impulso de actividades
econmicas de valor agregado, incluyendo la elaboracin y
exportacin de artesanas y el ecoturismo. Ms de cien familias
han mejorado sus ingresos entre 100 y 500% en los ltimos tres
aos gracias a la exportacin a EE. UU. de artesanas de la fbra
de la palmera chambira (PROCREL 2010)
Con base en la aplicacin de matrices de monitoreo comu-
nitario se ha podido determinar el dramtico descenso de las
amenazas para los recursos de la biodiversidad en las zonas donde
se implement el proyecto (Figs. 6 9, PROCREL 2010)
Tambin se ha comprobado una notable recuperacin de las
pesqueras por efectos del manejo comunitario (del orden del
40% en cinco aos), en especial en la Quebrada Yanayacu, rea
de amortiguamiento del rea de Conservacin Regional Comu-
nal Tamshiyacu-Tahuayo, en donde el trabajo con comunidades
es ms reciente y se ha monitoreado la captura por unidad de
esfuerzo desde el inicio de la aplicacin de las medidas de manejo
(Fig. 10; PROCREL 2011)
Conclusiones
Las comunidades locales pueden no ser el modelo perfecto de
gestin del bosque amaznico. Estn compuestas por personas,
y las personas cometen errores y tambin pueden llegar a ser
cortoplacistas y egostas; los indgenas y ribereos amaznicos
no son una excepcin, no existe el noble salvaje (Alvard 1993).
La llamada tragedia de los bienes comunes es actualmente
una de las ms graves amenazas para la salud de los ecosistemas
amaznicos: la falta de control del Estado y el colapso de las
sociedades indgenas favorecen la extraccin indiscriminada y
sin medidas de manejo de recursos de fora y fauna (Smith 2003;
Ostrom 2003). Y la tragedia de los bienes comunes tambin
puede ocurrir dentro de las comunidades, cuando no ejercen
control sobre las acciones de sus miembros.
Los resultados de los proyectos impulsados por el IIAP y algu-
nas otras organizaciones muestran que s se puede mejorar de la
calidad de vida y de la economa de las poblaciones amaznicas
al mismo tiempo que se revierten los procesos de prdida de
bosques y degradacin de hbitats, contribuyendo por tanto a
conservar de forma efectiva y sostenible la diversidad biolgica en
Figura 6. Nmero de infractores que ingresaron a extraer madera. Figura 7. Porcentaje de comuneros que realizan practicas destruc-
tivas para pescar.
Figura 8. Descenso de ingreso de infractores al rea de infuencia
del rea de Conservacin Regional Comunal Tamshiyacu-Tahuayo
(ACRACTT).
Figura 9. Porcentaje de comuneros que extraen palmeras para
techado con prcticas destructivas.
15
13
13,24
21,44
18,63
25,5
0
5
10
15
20
25
30
2005 2006 2007 2008 2009 2010
C
P
U
E
Figura 10. Captura de pescado por unidad de esfuerzo (nmero de
individuos capturados/nmero promedio de red-trampa utilizadas)
en la Quebrada Yanayacu, rea de Conservacin Regional Comunal
Tamshiyacu-Tahuayo (ACRACTT).
200
150
70
0
7
0 0 0 0 0 0 0
0
20
40
60
80
100
120
140
160
180
200
2006 2007 2008 2009
Aos
N
m
e
r
o
d
e
t
r
o
z
a
s
San Juan
Ayacucho
Esperanza
Cunshico
San Pedro
Diamante
Buena Vista
Chino
6 6
0 0
90
25
0
5
4
8
1
7
1
5
0
10
20
30
40
50
60
70
80
90
100
2006 2007 2008 2009
Aos
P
o
r
c
e
n
t
a
j
e
s
San Juan
Ayacucho
Esperanza
Cunshico
672
637
292
76
0
100
200
300
400
500
600
700
800
2006 2007 2008 2009
os
N
m
e
r
o
d
e
I
n
f
r
a
c
t
o
r
e
s
28
40
10
7
3 3 3
0 0 0 0
2
0 0 0 0 0
5
10
15
20
25
30
35
40
2006 2007 2008 2009
Aos
P
o
r
c
e
n
t
a
j
e
San Juan
Ayacucho
Esperanza
Cunshico
San Pedro
Diamante
Buena Vista
Chino
231
Gestin participativa de la biodiversidad con comunidades amaznicas
Rev. peru. biol. 19(2): 223 - 232 (August 2012)
grandes espacios naturales. No cabe duda de que las comunidades
pueden ser mucho mejores gestoras que el Estado, al menos en
el escenario de la Amazona baja del Per.
Sin embargo, es ms que claro que actualmente las comunida-
des amaznicas necesitan una buena dosis de acompaamiento
y capacitacin para que salgan de la vorgine extractivista en la
que estn metidos desde hace dos o tres generaciones: extraccin
descontrolada de recursos de fora y fauna - empobrecimiento
creciente de la poblacin - ms presin sobre los recursos. Es
un crculo vicioso que se agrava ao tras ao, a medida que se
articulan ms con el mercado y requieren ms bienes de consu-
mo (Trujillo 2009). Para garantizar la sostenibilidad econmica
del modelo de conservacin efectiva de grandes paisajes es
indispensable el desarrollo de alternativas productivas con arti-
culacin al mercado.
Un aspecto importante a tener en cuenta en estas experiencias
es que se ha obtenido avances signifcativos enfrentando dos de
los graves retos en la Amazona: impulsar el desarrollo sostenible
de las comunidades locales y promover la conservacin efectiva
de los bosques y otros ecosistemas. Cerca de dos millones de
hectreas estn ahora efectivamente protegidas en varias reas
protegidas de nivel nacional y regional en Loreto; en ellas se
recuperan ecosistemas y recursos sobre explotados en el pasado,
de fauna y fora, mientras que se aprovechan con medidas de
manejo recursos de fauna y productos forestales diferentes a la
madera para benefcio de las comunidades.
En las reas con mayor tiempo de gestin las comunidades
han mejorado tanto su calidad de vida (dada la recuperacin de la
fauna y la fora que constituyen la base de su economa, y gracias
al involucramiento en actividades econmicas de agregacin de
valor y comercializacin de productos de la biodiversidad) que
se han convertido en un modelo de desarrollo para otras reas y
para otras regiones. El Gobierno Regional de Loreto ha asumido
al PROCREL y sus lneas estratgicas como un proyecto emble-
mtico en el marco de su visin de Regin Productiva; y varios
pueblos indgenas han mostrado inters en replicar el modelo
de co-gestin comunal y conservacin productiva en sus zonas,
mientras que en la Regin Ucayali se est ejecutando un proyecto
similar al de Loreto. Las regiones Madre de Dios y Amazonas,
por su parte, han mostrado tambin inters en replicar el modelo.
Literatura citada
Alvard M. S. 1993. Testing the Ecologically Noble Savage
Hypothesis: Interspectifc prey choice by Piro Hunters
of Amazonian Peru. Human Ecology 21(4): 355 387.
Alvarez J. 2006. Gestin comunal y territorio: lecciones aprendidas
de la cuenca del Nanay (Amazona norperuana) para el
manejo de la fauna silvestre amaznica. Revista electrnica
Manejo de fauna silvestre en la Amazona. N 1, Vo. 1.
lvarez J. 2007. Comunidades locales, conservacin de la avifauna
y de la biodiversidad en la Amazona peruana. Rev. Peru.
Biol. 14(1): 151-158.
lvarez J., F. Rojas M. y A. J. Araujo. 2004. Una experiencia de
manejo comunal de los recursos del bosque en la Reserva
Nacional Allpahuayo - Mishana. Documento Tcnico
N 11, BIODAMAZ, IIAP. http://www.iiap.org.pe/
biodamaz/faseii/download/literatura_gris/Articulos%20
cient%C3%ADfcos/ [Acces 03.07.2011].
Anderson. A. (Ed.) 1989. Alternatives to Deforestation Steps Toward
Sustainable Uses of Amazonian Forests. Columbia Uni-
versity Press, New York.
Asner G.P., G.V.N. Powell, J. Mascaro, et al. 2003. Who should pay
for tropical conservation and how could the costs be met?
Oryx 37: 238250.
Balmford A. & T. Whitten 2003. Who should pay for tropical conser-
vation and how could the costs be met? Oryx 37: 238250.
Bodmer R. 2000. Integrating hunting and protected areas in the
Amazon. Pp. 277-290 en N. Dunstone and A. Entwistle
(Eds.) Future Priorties for the Conservation of Mammals:
Has the Panda had its Day? Cambridge University Press,
Cambridge, UK.
Bodmer R. & P. Puertas 2000. Community-based comanagement
of wildlife in the Peruvian Amazon. Pp. 395-409 en J.
Robinson and E. Bennett (Eds.) Hunting for Sustainability
in Tropical Forests. Columbia University Press, New York.
Bodmer R.E., J.W. Penn, P.E. Puertas, L. Moya I. & T.G. Fang. 1997.
Linking conservation and local people through sustainable
use of natural resources: Community-based management
in the Peruvian Amazon. Pp. 315-358 en C. Freese (Ed.)
Harvesting Wild Species. John Hopkins University Press.
Brown K. 2003. Three challenges for a real people-centred con-
servation. Global Ecology and Biogeography, 12, 8992.
Bruner A.G., R.E. Gullison, R.E Rice & G.A.B. da Foncesca. 2001.
Effectiveness of parks in protecting tropical biodiversity.
Science 291: 125128.
Butler R.A. & W.F. Laurance. 2009. Is oil palm the next emerging
threat to the Amazon? Tropical Conservation Science
Vol. 2(1):1-10.
CADMA. 1992. Amazona sin mitos. Comisin Andina de Desa-
rrollo y Medio Ambiente. Ed. Banco Interamericano de
Desarrollo/PNUD/TCA.
Campbell L.M. & A. Vainio-Mattilia. 2003. Participatory develo-
pment and community based conservation: opportunities
missed for lessons learned. Human Ecology, 31, 417437.
CIAM. 2011. La Amazona vale un Per. Consejo Interregional
Amaznico CIAM, documento tcnico. Lima.
Clad J. 1984. Conservation and indigenous peoples: a study of
convergent interests. Cultural Survival Quarterly 8: 6873.
Clay J. 1988. Indigenous Peoples and Tropical Forests. Models of
Land Use and Management from Latin America. Cultural
Survival. Inc. Cambridge, MA. Cultural Survival Report
27: 69-73.
CSD - Commission on Sustainable Development (2004) Partners-
hips for Sustainable Development: Report of the Secre-
tary General. Http://www.un.org/esa/sustdev/csd/csd12/
csd12_docs.htm [Acces. 23.12.2010].
Cooke B. & U. Kothari (Eds.) 2001. Participation: The New Tyranny?
Zed Books, London, UK, 207 pp.
Danielsen F., M. M. Mendoza, A. Tagtag, et al. 2007. Increasing
Conservation Management Action by Involving Local
People in Natural Resource Monitoring. Ambio 36, 7:
566-570.Davis S. H. and A. Wali. 1994. Indigenous Land
Tenure and Tropical Forest Management in Latin America.
Ambio Vol. 23 No. 8.
Davis S. H. & A. Wali. 1994. Indigenous Land Tenure and Tropical
Forest Management in Latin America. Ambio Vol. 23 No. 8.
De Soto H. 2010. La Amazona no es Avatar. El Comercio, Per
05.06.2010.
Ferraro P. J. & S. K Pattanayak. 2006. Money for nothing? A call
for empirical evaluation of biodiversity conservation
investments. PLoS Biology, 4, e105.
Garca Villacorta R. 2010. Identifcacin de Procesos Ecolgicos
y Evolutivos Esenciales para la Conservacin de la
Biodiversidad en la Regin Loreto. Documento Tcnico
Proyecto Apoyo al Programa de Conservacin, Gestin y
Uso Sostenible de la Biodiversidad en la Regin Loreto.
122 pp. Iquitos.
232
lvarez & Shany
Rev. peru. biol. 19(2): 223 - 232 (Agosto 2012)
Gasch J. 1999. Desarrollo rural y pueblos indgenas amaznicos.
Ed. Abya-Yala. Quito.
Gasch J. 2004: Una concepcin alternativa y crtica para pro-
yectos de desarrollo en la Amazona. Pp. 105-118 en J.
Gasch (Ed.): Crtica de proyectos y proyectos crticos
de desarrollo. Iquitos, Instituto de Investigaciones de la
Amazona Peruana.
Gasch J. 2010. Conferencia en la Universidad Nacional de Colom-
bia. IIAP, manuscrito, 25 pp
Getz W.M., Fortmann, L., Cumming, D., et al. (Eds.) 1997. Social
Change and Conservation: Environmental Politics and
Impacts of National Parks and Protected Areas. Earthscan,
London, UK.
Ghimire, K.B. & M.P. Pimbert (Eds.) 1997. Social Change and Con-
servation: Environmental Politics and Impacts of National
Parks and Protected Areas. Earthscan, London, UK.
Goodman D. & A. Hall. A. (Eds.) 1990. The Future of Amazonia:
Destruction or Sustainable Development. St. Martin's
Press, New York.
Guha R. 1997. The authoritarian biologists and the arrogance of
anti-humanism: wildlife conservation in the third world.
The Ecologist 27: 1420.
Holt F. L. 2005. The catch-22 of conservation: indigenous peoples, bio-
logists and cultural change. Human Ecology 33: 199215.
Houtart F. 2011. El escndalo de los agrocombustibles en los pases
del Sur http://alainet.org/active/47497. [acces. 10.06.11].
IIAP. 2004. Informe fnal del Proyecto Conservacin de la biodi-
versidad y manejo comunal de los recursos naturales en la
cuenca del ro Nanay Banco Mundial/GEF. Documento
Tcnico, Instituto de Investigaciones de la Amazona
Peruana, Iquitos.
Johnson A. 1989. How the Machiguenga manage resources: conser-
vation or exploitation of nature? Advances in Economic
Botany 7: 213222.
Novaro A., Redford, K. & R. Bodmer. 2000. Effect of hunting
in source-sink systems in the Neotropics. Conservation
Biology 14(3): 713721.
Ostrom E. 2003. Reformulando los bienes comunes. Pp. 48-77 en
Smith R. Ch. & D. Pinedo (Eds.). El cuidado de los bienes
comunes. Gobierno y manejo de los lagos y los bosques
en la Amazona. Lima, Ed. IEP.
Parry L., C. A. Peres, B. Day & S. Amaral. 2010. Ruralurban
migration brings conservation threats and opportunities
to Amazonian watersheds. Conservation Letters xx 19.
Peres C. 1994. Indigenous reserves and nature conservation in
Amazonian forests. Conservation Biology 8(2): 586-588.
Peres C. 2000. Evaluating the impact and sustainability of sub-
sistence hunting at multiple Amazonian forest sites. Pp.
31-56 en J. Robinson and E. Bennett (Eds.) Hunting for
Sustainability in Tropical Forests. Columbia University
Press, New York.
Phillips O., G. Lpez-Gonzlez, Y. Malhi, et al. 2009. Drought Sen-
sitivity of the Amazon Rainforest. Science 323: 1344-1347.
Porter-Bolland L., E. A. Ellis, M. R. Guariguata, et al. 2011. Com-
munity managed forests and forest protected areas: An
assessment of their conservation effectiveness across
the tropics. Forest Ecol. Manage. doi:10.1016/j.fore-
co.2011.05.034
Posey D. & B. Balee (Eds.) 1989. Resource Management in Amazo-
nia: Indigenous and Folk Strategies. New York Botanical
Garden. New York. Advances in Economic Botany. Vol. 7.
PROCREL 2011. Informe de monitoreo participativo de la pesca en
la Quebrada Yanayacu, zona de amortiguamiento del rea
de Conservacin Regional Comunal Tamshiyacu-Tahuayo.
Informe Tcnico Programa de Conservacin, Gestin y
Uso Sostenible de la Biodiversidad en la Regin Loreto
PROCREL. Iquitos.
PROCREL. 2010. Informe de monitoreo participativo en la zona
de amortiguamiento del rea de Conservacin Regional
Comunal Tamshiyacu-Tahuayo. Informe Tcnico Pro-
grama de Conservacin, Gestin y Uso Sostenible de la
Biodiversidad en la Regin Loreto PROCREL. Iquitos.
Pyhl Aili. 2003. Productive Conservation in Amazonia: Institu-
tions, Participation and Markets. PhD Thesis submitted
to the School of Development Studies, University of East
Anglia, U.K.
Redford KH. & A.M. Stearman. 1993. Forest-dwelling native Ama-
zonians and the conservation of biodiversity: interests in
common or in collision? Conservation Biology 2: 248255.
Redford K.H. 1992. The empty forest. Bioscience 42: 412422.
Redford K.H. & S. Taber. 2000. Writing the wrongs: developing a
safe-fail culture in conservation. Conservation Biology
14: 15671568.
Roe D. & M. Jack. 2001. Stories from Eden: Case Studies of
Community-based Wildlife Management. Evaluating
Eden Series 9. International Institute for Environment and
Development, London, UK.
Sanderson S. E. & K. H. Redford. 2003. Contested relationships
between biodiversity conservation and poverty alleviation.
Oryx 37: 389390.
Santoyo A. 2004. Representaciones y programa de cambio en la
Amazona durante el siglo XIX: Anlisis del pensamiento
civilizador colombiano. Pp. 13-28 en J. Gasch (Ed.) Crti-
ca de proyectos y proyectos crticos de desarrollo. Iquitos,
Instituto de Investigaciones de la Amazona Peruana.
Schwartzman S. & B. Zimmerman. 2005. Conservation alliances
with indigenous peoples of the Amazon. Conservation
Biology 19: 721727.
Sheil D. & A. Lawrence. 2004. Tropical biologists, local people and
conservation: new opportunities for collaboration. Trends
Ecol. Evol. 19, 634638.
Smith D Ch. 2003. Los bienes comunes y su gestin comunitaria:
conceptos y prcticas. Pp. 13-30 en D. Ch. Smith & D.
Pinedo (Eds.). El cuidado de los bienes comunes. Gobier-
no y manejo de los lagos y los bosques en la Amazona.
Lima, Ed. IEP.
Smolker R., B. Tokar., A. Petermann, E. Hernndez. 2007. El verda-
dero costo de los agrocombustibles: alimentacin, bosques
y clima. Global Forest Coalition, 80 pp.
Terborgh J. 1999. Requiem for Nature. Island Press, Washington,
DC, USA.
Terborgh J., G. Nuez-Iturri, N. C. A. Pitman, et al. 2008. Tree
recruitment in an empty forest. Ecology, 89(6): 17571768
Trujillo Osorio C. 2009. Efectos de la integracin al mercado sobre
las formas de uso del bosque tropical en territorios ind-
genas del Amazonas. Tesis para acceder a la Maestra en
Estudios Amaznicos de la Univ. Nacional de Amazonia
(Leticia, Colombia).
Vandermeer J. & I. Perfecto. 2007a. Tropical conservation and grass
roots social movements: Ecological theory and social
justice. Bulletin of the Ecological Society of America
88:171-175.
Vandermeer J. & I. Perfecto. 2007b. The agricultural matrix and a
future paradigm for conservation. Conservation Biology
21:274277.
Vermeulen S. & D. Sheil. 2007. Partnerships for tropical conserva-
tion. Oryx 41(4): 434440.
Wright S. J. & H.C. Muller-Landau. 2006. The Future of Tropical
Forest Species. BIOTROPICA 38(3): 287301.
Wright A. & W. Wolford. 2003. To inherit the earth: the landless
movement and the struggle for a new Brazil. Food First
Books, Institute for Food and Development Policy, San
Francisco, California, USA.
233
Rev. peru. biol. 17(2): 000- 000 (August 2010)
Colofn
234
Suscripciones y Canje
Subscriptions and Exchange programs
La Revista Peruana de Biologa es publicada en abril. agosto y diciembre y esta dedicada a la publicacin de los resultados de
investigaciones originales e inditas en las reas de Biodiversidad, Biotecnologa, Manejo ambiental, Ecologa y Biomdicas. Los
trabajos recibidos son evaluados por rbitros segn criterios internacionales de calidad, creatividad, originalidad y contribucin
al conocimiento.
Te Revista Peruana de Biologa is published in April, August and December, the aims is to disseminate the results of original
research in Biodiversity, Biotechnology, Environmental management, Ecology and Biomedical areas. Papers in Spanish or English are
peer-reviewed using international criteria of quality, creativity, originality and the knowledge contribution.
Revista Peruana de Biologa
Suscripcin anual, costo incluye envo.
Annual subscription, mailing is included.
Per 300 nuevos soles
Other countries US$ 250
Nombre/name:..
Direccin postal/Full postal address:
Favor de extender Cheque certifcado u orden de pago a nombre de la Facultad de Ciencias Biolgicas-UNMSM.
Please make check or money order payable to Facultad de Ciencias Biolgicas-UNMSM.
Enviar a:
Send to:
Leonardo Romero
Editor, Revista Peruana de Biologa
Facultad de Ciencias Biolgicas-UNMSM
Casilla Postal: 11-0058
Lima-11,
Per
Mayor informacin dirigirse a:
For futher information contact:
Editor Jefe, Leonardo Romero
Telfono (511) 619-7000-1502/ Telefax (511) 619-7000-1509
email: editor.revperubiol@gmail.com
235
Rev. peru. biol. 17(2): 000- 000 (August 2010)
PAUTAS PARA LA PRESENTACIN DE LOS ARTCULOS EN LA REVISTA PERUANA DE BIOLOGA
Abril 2012
(Contina....)
IDENTIDAD Y PROPSITO
La Revista Peruana De Biologa es una publicacin cientfca arbitrada producida por el Instituto de Investigaciones de
Ciencias Biolgicas Antonio Raimondi, Facultad de Ciencias Biolgicas, Universidad Nacional Mayor de San Marcos, Lima, Per;
es publicada tres veces al ao y los nmeros aparecen en abril, agosto y diciembre, tanto en su versin impresa como Online.
La Revista Peruana De Biologa publica artculos completos, originales e inditos que contribuyan al conocimiento cient-
fco en los temas de biodiversidad, biotecnologa, ecologa, manejo ambiental y biomedicina elaborados segn las normas
indicadas en las presentes pautas.
EVALUACIN DE LOS TRABAJOS
Los trabajos que cumplan con las pautas solicitadas sern incluidos en el la lista para evaluacin, un editor ser encargado de
conducir el proceso envindo el trabajo a rbitros. El trabajo ser evaluado segn criterios internacionales de calidad, crea-
tividad, originalidad y contribucin al conocimiento. El artculo ser aceptado luego del proceso de revisin por rbitros y
de realizadas las modifcaciones indicadas. El artculo aceptado ser editado y una prueba enviada al autor para la aceptacin
y consentimiento de publicacin.
CARTA DE PRESENTACIN
El trabajo ser acompaado de mensaje email dirigido al Editor Jefe, indicando haber ledo las presentes pautas para presenta-
cin de trabajos, asegurando la originalidad, carcter indito y completo del trabajo presentado y su disposicin para que
sea revisado y editado. Esta carta tendr carcter de declaracin jurada segn lo estipulan las directivas del Vicerrectorado
de Investigacin de la Universidad Nacional Mayor de San Marcos referidos a las autoras de trabajos publicados. Modif-
caciones en las autoras sern motivo para detener el proceso de edicin. Las coordinaciones se llevaran entre el Editor Jefe
y el responsable del trabajo.
PRESENTACIN DE LOS TRABAJOS
Los archivos del trabajo sern enviados por email al correo del editor: editor.revperubiol@gmail.com
Los trabajos pueden ser presentados en idioma ingls o castellano.
El trabajo debe tener tres partes bsicas: (a) identifcacin, (b) cuerpo y (c) literatura citada
(a) La identificacin debe contener: Ttulo (en ingls y castellano), nombre y apellido de los autores, nombre completo de la
institucin de los autores, correo electrnico de cada uno de los autores, y una direccin postal del autor para correspondencia.
Resumen no mayor de 250 palabras (en ingls y castellano), 5 palabras clave (en ingls y castellano).
(b) El cuerpo del trabajo variar segn la seccin de la Revista:
1. TRABAJOS ORIGINALES. Son artculos primarios, inditos que exponen los resultados de trabajos de investigacin y
constituyen aportes al conocimiento. Se incluyen aqu la descripcin de especies nuevas, que cumplan con las Normas
y caractersticas de la informacin en los trabajos (ver abajo). El cuerpo est organizado en: Introduccin, Material y
mtodos, Resultados, Discusin y Agradecimientos, debe abarcar un texto promedio de 20 pginas, las ilustraciones (Tablas
y Figuras) deben ser slo las necesarias para una mejor exposicin de los resultados.
2. NOTAS CIENTFICAS. Son artculos primarios, se incluyen aqu: reportes de distribucin, inventarios taxonmicos, notas
taxonmicas, resultados de ensayos de laboratorio, entre otros y cuya informacin es de inters para la comunidad cient-
fca. La extensin del texto no ser mayor de 8 pginas. Esta seccin puede estar organizada como: cuerpo de la Nota (en
algunos casos: introduccin, material y mtodos, resultados y discusin) y Agradecimientos.
3. COMENTARIOS. Son artculos donde se discute y exponen temas o conceptos de inters para la comunidad cientfca. Se
incluyen aqu ensayos de opinin y monografas. Deben contar con las siguientes partes: cuerpo del comentario y Agrade-
cimientos. Todo el artculo debe tener un texto promedio de 10 pginas.
4. COMENTARIOS DE LIBROS. Son artculos que comentan recientes publicaciones de inters para la comunidad cientfca.
Puede solicitarse al Comit Editor la elaboracin de un comentario enviando dos copias del libro a la direccin postal de
la Revista Peruana de Biologa.
Los Agradecimientos. Se indicara la fuente de fnanciamiento y deben dedicarse solamente a las personas e instituciones
que colaboraron directamente en la realizacin del trabajo.
Informacin adicional. Debe incluirse despus de Agradecimientos debe incluirse informacin sobre la contribucin
de cada uno de los autores (revisar http://www.icmje.org/ethical_1author.html) y de no incurrir en conficto de intereses
(revisar http://www.icmje.org/ethical_4conficts.html).
(c) Citacin y Literatura Citada. Las CITAS EN EL TEXTO deben incluir el apellido del autor y ao sin comas que los separen
(ejemplo: (Carrillo 1988) o ... de acuerdo a Snchez (1976) Otros ejemplos:
la concentracin de nitrgeno fue medida segn Chvez y Castro (1998).
de nitrgeno se midi por el mtodo colorimtrico (Chvez & Castro 1998).
la diversidad de especies es considerada elevada (Simoniz 2010, Perez 1999, Aquino 1988).
Si hay varios trabajos de un autor en un mismo ao, se citar con una letra en secuencia adosada al ao (ejemplo: Castro 1952a).
Cuando hay ms de dos autores se citar al primer autor y se colocar et al. (Ejemplo: (Smith et al. 1981) o segn Smith
et al. (1981)).
La LITERATURA CITADA incluir todas las referencias citadas en el texto dispuestas solamente en orden alfabtico y sin numeracin.
La cita se inicia con el apellido del primer autor a continuacin, sin coma, las iniciales del nombre separadas con puntos y
236
sin espacio. El segundo y tercer autor deben de tener las iniciales de los nombre y a continuacin el apellido. El ltimo autor
se diferenciara por que le antecede el smbolo &. Si hubiesen ms de tres autores pueden ser indicados con la abreviatura et
al. En la literatura citada solamente se usa letra tipo normal, no itlica, no versalita. Ejemplos:
Montgomery G.G., R.C. Best & M. Yamakoshi. 1981. A radio-tracking study of the American manatee Trichechus
inunguis (Mammalia: Sirenia). Biotropica 13: 81 -85.
Buhrnheim C.M. & L.R. Malabarba. 2006. Redescription of the type species of Odontostilbe Cope, 1870 (Teleostei:
Characidae: Cheirodontinae), and description of three new species from the Amazon basin. Neotrop. ichthyol. 4 (2):
167-196.
Nogueira R.M.R., M.P. Miagostovich, H. G. Schatzmayr, et al. 1995. Dengue type 2 outbreak in the south of the
State of Bahia, Brazil: laboratorial and epidemiological studies. Rev. Inst. Med. trop. S. Paulo 37 (6): 507-510.
McLachlan A. & A.C Brown. 2006. Te Ecology of Sandy Shores. Elsevier Science & Technology Books. 373pp.
Crawford D.J. 1983. Phylogenetic and systematic inferences from electrophoretic studies. In: S.D. Tanksley and T.J.
Orton, eds. Isozymes in plant genetics and breeding, Part A. Elsevier, Amsterdam. Pp. 257-287.
Pianka E.R. 1978. Evolutionary ecology. 2nd edn. New York: Harper & Row.
Carroll S.B. 2005. Evolution at Two Levels: On Genes and Form. PLoS Biol 3(7): e245. <http://biology. plosjournals.
org/archive/1545-7885/3/7/pdf /10.1371_journal.pbio.0030245-S.pdf >. Acceso 31/07/2005.
Food and Drug Administrations (FDA). 2001. Fish and Fishery Products Hazards and Controls Guidance. Tird
Edition June 2001. <http://www.cfsan.fda.gov/~comm/haccp4.html> (acceso 24/12/07).
CONAM. 2005. (en lnea). Informe nacional del estado del ambiente 2001. <http://www.conam.gob.pe /sinia/
INEA2001.shtml>. Acceso 31/07/2005.
IMARPE. 2002. (en lnea). Segundo informe del BIC Jos Olaya Balandra. Paita Salaverry. 24 febrero- 05 Marzo
2002. <http://www.imarpe.gob.pe /imarpe/informeolaya02-032002.php>. Acceso 01/07/2005.
Solari S.A. 2002. Sistemtica de Tylamys (mammalia: didelphimorphia: marmosidae). Un estudio de las poblacio-
nes asignadas a Tylamys elegans en Per. Tesis, Magster en Zoologa, mencin Sistemtica y Evolucin. Facultad de
Ciencias Biolgicas Universidad Nacional Mayor de San Marcos. <http://www.cybertesis.edu.pe/sisbib/2002/solari_ts /
html/index-frames.html>. Acceso 31/07/2005
Las citas de artculos en prensa deben incluir el volumen, el ao y el nombre de la revista donde saldrn publicados; de lo
contrario debern ser omitidos.
No se aceptan las citas a resmenes de eventos acadmicos (congresos y otros).
NORMAS Y CARACTERSTICAS DE LA INFORMACIN EN LOS TRABAJOS
La Revista Peruana de Biologa se gua de los conceptos ticos de autora mencionados en los Uniform Requirements for Ma-
nuscripts Submitted to Biomedical Journals (http://www.icmje.org/), asumimos la veracidad de la carta de presentacin y
de la carta de consentimiento de publicacin.
Cuando el artculo exponga sobre experimentos con humanos y animales, los procedimientos deben de ceirse a la Declaracin
de Helsinki de 1975 y a las leyes peruanas vigentes (Ley 27265). Deben ser presentadas las declaraciones pertinentes y
mencionadas en el texto.
Cuando el artculo exponga sobre nuevas especies, nuevos registros, ampliaciones biogeogrfcas o inventarios taxonmicos debe
tomarse en cuenta la informacin requerida en el Darwin Core. Tambin debe indicarse el depsito de los ejemplares en un
centro de referencia taxonmico (v.g. Museo).
Las referencias a localidades y puntos de colecta deben ser referenciados geogrfcamente.
Cuando los especmenes hayan sido colectados en reas protegidas, debe de indicarse los respectivos permisos.
Los nombres cientfcos del gnero y especie irn en cursivas. La primera vez que se cita un organismo deber hacerse con su
nombre cientfco completo (gnero, especie y autor); posteriormente podr citarse solamente la inicial del nombre genrico
y el nombre especfco completo. Para el caso de las abreviaturas de autores en nombres botnicos pueden referirse a la base
de datos TROPICOS (http://www.tropicos.org/) y AlgaeBase (http://www.algaebase.org/).
Deben usarse los smbolos de las unidades del Sistema Internacional de Medidas. Si fuera necesario agregar medidas en otros
sistemas, las abreviaturas correspondientes deben ser defnidas en el texto.
Los textos en castellano debern utilizar la coma decimal, no punto (ejemplo correcto: 0,5; incorrecto: 0.5). Los textos en ingles
deben usar punto decimal.
ILUSTRACIONES
Las Figuras (mapas, esquemas, diagramas, dibujos, grfcos, fotos, etc.) sern numeradas correlativamente con nmeros arbigos;
de igual manera las Tablas. Las leyendas de las fguras y Tablas deben presentarse a continuacin del texto y ser sufciente-
mente explicativas.
Las fguras escaneadas deben guardarse en un archivo TIFF, tamao del original, 300 dpi. Las grfcas deben de enviarse en for-
mato nativo editable (achivo.xls, archivo.wmf, archivo.svg, archivo.eps), no como imgenes (JPGE, TIFF, PNG). Los mapas
pueden enviarse en formatos SHP. Fotos de cmaras digitales en formato JPGE mayor a 3 Mpixel. Otros archivos de imgenes
en TIFF, BMP, JPGE de alta resolucin y tamao, y fguras vectoriales en Ai, PSD. Formatos procedentes de software como
R, Statistica, en formatos editables de PDF, EPS (Postscript). Costos por ilustraciones a color sern asumidos por el autor.
(Contina....)
237
Rev. peru. biol. 17(2): 000- 000 (August 2010)
Los archivos deben presentarse por separado, esto es, un archivo con el texto y leyendas en formato MS-Word. Otro archivo
para las tablas en MS-Excel o como tablas en MS-Word. Otros archivos en formatos nativos, no como imgenes insertadas
en otros archivos (por ejemplo no enviar imgenes pegadas en una hoja de MS-Word o Excel).
CONSENTIMIENTO DE PUBLICACIN
El autor responsable recibir una prueba del trabajo en formato PDF, el cual deber revisarlo cuidadosamente, y consultarlo con los
otros autores. Una vez solucionados errores de edicin, el autor responsable enviara una carta o email indicando que el trabajo fue
revisado por todos los autores y estn conformes y mencionar su consentimiento de para la publicacin del trabajo como artculos
de la Revista Peruana de Biologa. Este es un requisito para la publicacin del trabajo. Con la prueba el autor responsable recibir el
COSTO POR PUBLICACIN.
COSTO POR PUBLICACIN.
El costo por publicacin se refere a gastos extras por manipulacin, edicin extraordinaria (v.g. elaboracin de ilustraciones, traducciones,
etc.) y por impresin de ilustraciones a color. El pago por publicacin se realiza en las ofcinas de la Facultad de Ciencias Biolgicas
de la Universidad Nacional Mayor de San Marcos, o en coordinacin con el Editor Jefe.
El autor principal podr solicitar cuatro ejemplares de la revista en las ofcinas de la Facultad de Ciencias Biolgicas de la Universidad
Nacional Mayor de San Marcos.
Comit Editor
Email: editor.revperubiol@gmail.com
Correo postal:
Leonardo Romero (Editor)
Revista Peruana de Biologa
UNMSM-FCB
Apartado 11-0058
Lima 11
Per
238
Revista PeRuana de Biologa
Volumen 19 Agosto, 2012 Nmero 2
Rev. peru. biol. ISSN 1561-0837
Contenido
HOMENAJE
127 A la memoria del doctor Abundio Sagstegui Alva (1932 2012)
In memoriam of Dr. Abundio Sagstegui Alva (1932 2012)
Eric F. Rodrguez Rodrguez y Blanca Len
Editorial
135 La Revista basurero
Te Journal landfll
Leonardo Romero
Trabajos originales
139 Una nueva especie de Salvia (Lamiaceae) del Norte de Per
A new species of Salvia (Lamiaceae) from Northern Peru
Abundio Sagstegui Alva y Eric F. Rodrguez Rodrguez
143 Una nueva especie de Deprea Raf. (Solanaceae) del Per
A new species of Deprea Raf. (Solanaceae) from Peru
Marco Antonio Cueva
e talo Francisco Trevio
149 Flora vascular de las lomas de Villa Mara y Amancaes, Lima, Per
Vascular fora from Villa Mara and Amancaes Lomas, Lima, Peru
Huber Trinidad, Elluz Huamn-Melo, Amalia Delgado y Asuncin Cano
159 Dieta de murcilagos flostmidos del valle de Kosipata, San Pedro, Cusco - Per
Diet of phyllostomid bats of Kosipata Valley, San Pedro, Cusco - Peru
Rossana Maguia, Jessica Amanzo y Luis Huamn
167 Reproduccin y alimentacin de Phytotoma raimondii, cortarrama peruana en El Gramadal, Ancash
Reproduction and food habits and of Phytotoma raimondii, Peruvian Plantcutter in El Gramadal, Ancash
Mario Rosina y Mnica Romo
175 Estructura de tallas de tortuga pico de loro Lepidochelys olivacea (Testudines: Cheloniidae) en Tumbes, Per
Size structure of Olive Ridley turtle Lepidochelys olivacea (Testudines: Cheloniidae) in Tumbes, Peru
Manuel Vera y Carlos A. Rosales
181 Patgenos introducidos al Per en post larvas de Litopenaeus vannamei importadas
Pathogen introduced to Peru by trade in postlarvae of Litopenaeus vannamei
Mervin Guevara y Rubn Alfaro
187 Optimizacin de la biorremediacin en relaves de cianuracin adicionando nutrientes y microorganismos
Optimization of cyanidation tailings bioremediation adding nutrients and microorganisms
Jasmin Hurtado y Arturo Berastain
193 Dosis nica de ciclofosfamida disminuye la calidad espermtica y el epitelio germinal masculino en ratones
Unique cyclophosphamide doses in male mice decrease the spermatic quality and germinal epithelium
Lyonal Acosta, Vctor Nez, Jonathan Vsquez, Jos Pino y Betty Shiga
199 Chemical composition of Apodanthera bifora, a Cucurbit of the dry forest in northwestern Peru
Composicin qumica de Apodanthera bifora, una cucurbitcea del bosque seco del noroeste peruano
Daniel Clark, Maribel Tupa, Andrea Bazn, Lily Chang and Wilfredo L. Gonzles
205 Diversidad gentica de papa nativa (Solanum spp.) conservada in situ en cinco regiones de Per
Genetic diversity of in situ conservation landraces potato (Solanum spp.) from fve regions of Peru
Julin Soto, Tulio Medina, Yeny Aquino y Rolando Estrada
Notas cientfcas
213 Descripcin del nido de Scolomys melanops (Rodentia, Cricetidae) y su relacin con Lepidocaryum tenue (Arecales, Arecaceae)
Description of Scolomys melanops nest (Rodentia, Cricetidae) and its relationship with Lepidocaryum tenue (Arecales, Arecaceae)
Edgardo M. Rengifo y Rolando Aquino
217 Ampliacin de la distribucin de Sephanoides sephaniodes (Trochilidae: Aves) en Argentina
Expansion of the distribution of Sephanoides sephaniodes (Trochilidae: Aves) in Argentina
Nancy Vernica Marinero, Ricardo Omar Cortez, Eduardo Alfredo Sanabria, Lorena Beatriz Quiroga
219 Herbarium Areqvipense (HUSA): informatizacin y representatividad de su coleccin
Herbarium Areqvipense (HUSA): computerization and representativeness of its collection
Italo F. Trevio, Diego A. Sotomayor, Marco A. Cueva, Rafael Perez, Laura Cceres, Daniel Ramos, Edgardo M. Ortiz y Victor Quipuscoa
Comentario
223 Una experiencia de gestin participativa de la biodiversidad con comunidades amaznicas
An experience on participatory management of biodiversity with Amazonian communities
Jos lvarez y Noam Shany
También podría gustarte
- Introduced Dung Beetles in Australia: A Pocket Field GuideDe EverandIntroduced Dung Beetles in Australia: A Pocket Field GuideAún no hay calificaciones
- Ex Situ Plant Conservation: Supporting Species Survival In The WildDe EverandEx Situ Plant Conservation: Supporting Species Survival In The WildEdward O. GuerrantAún no hay calificaciones
- RPB V16n1-Mach PDFDocumento146 páginasRPB V16n1-Mach PDFLeonardo Romero0% (2)
- RPB v19n1 MachoteDocumento134 páginasRPB v19n1 MachoteLeonardo RomeroAún no hay calificaciones
- RPB v15n1 MachDocumento152 páginasRPB v15n1 MachLeonardo RomeroAún no hay calificaciones
- AB-WA-2013 WA Australasian Bittern Project Report PDFDocumento122 páginasAB-WA-2013 WA Australasian Bittern Project Report PDFEguene SofteAún no hay calificaciones
- Kakadu RegionDocumento209 páginasKakadu RegionD.s.LAún no hay calificaciones
- Plantindicatorsr00clemuoft BWDocumento602 páginasPlantindicatorsr00clemuoft BWMichaiel PiticarAún no hay calificaciones
- Gunther Kunkel (Auth.) Flowering Trees in Subtropical Gardens 1978Documento342 páginasGunther Kunkel (Auth.) Flowering Trees in Subtropical Gardens 1978Ignacio Garcia Ruiz100% (1)
- Crop Tree Field Guide - Selecting and Managing Crop Trees in The Central Appalachians (USDA, 2001)Documento140 páginasCrop Tree Field Guide - Selecting and Managing Crop Trees in The Central Appalachians (USDA, 2001)jonowelserAún no hay calificaciones
- GWW Final Report PDFDocumento252 páginasGWW Final Report PDFEguene SofteAún no hay calificaciones
- Global Survey of Ex Situ Conifer CollectionsDocumento50 páginasGlobal Survey of Ex Situ Conifer CollectionsChromaticghost2Aún no hay calificaciones
- Flora of Peru 31 Fi MacbDocumento530 páginasFlora of Peru 31 Fi MacbFrancisco DE Asis100% (1)
- Oxalis Del PERUDocumento290 páginasOxalis Del PERUpanchofrancoAún no hay calificaciones
- Proceedings of Gen 98 Sci eDocumento1274 páginasProceedings of Gen 98 Sci ejonho2006Aún no hay calificaciones
- Taxa of Vascular Plants Endemic To The Carpathian MTS: Preslia March 2016Documento59 páginasTaxa of Vascular Plants Endemic To The Carpathian MTS: Preslia March 2016cntudoseAún no hay calificaciones
- Guides To The Freshwater Invertebrates of Southern Africa Volume 5 - Non-ArthropodsDocumento304 páginasGuides To The Freshwater Invertebrates of Southern Africa Volume 5 - Non-ArthropodsdaggaboomAún no hay calificaciones
- Water Garden em Bra 01 TricDocumento154 páginasWater Garden em Bra 01 Tricliketoread100% (1)
- 1554Documento198 páginas1554grjhbAún no hay calificaciones
- Cambodia National Report On Protected Areas and DevelopmentDocumento150 páginasCambodia National Report On Protected Areas and Developmentyeangdonal100% (1)
- GTR Se020Documento228 páginasGTR Se020AxyAún no hay calificaciones
- (2005) Tropical Forests of The Guiana Shield Ancient Forests in A Modern World PDFDocumento541 páginas(2005) Tropical Forests of The Guiana Shield Ancient Forests in A Modern World PDFBruna BritoAún no hay calificaciones
- Checklist of Vascular Plants of The Southern Rocky Mountain RegionDocumento316 páginasChecklist of Vascular Plants of The Southern Rocky Mountain Regionfriends of the Native Plant Society of New MexicoAún no hay calificaciones
- Sanskrit Manuscripts in Raghunath Temple Library - M.A. Stein - Part2Documento177 páginasSanskrit Manuscripts in Raghunath Temple Library - M.A. Stein - Part2Dharmartha Trust Temple Library, JammuAún no hay calificaciones
- ManualDocumento83 páginasManualQky Rizky WulanAún no hay calificaciones
- Flora of British in 02 Hook RichDocumento808 páginasFlora of British in 02 Hook Richsksingl350100% (1)
- Promoting The Conservation and Use of Under Utilized and Neglected Crops. 08 - ChayoteDocumento58 páginasPromoting The Conservation and Use of Under Utilized and Neglected Crops. 08 - ChayoteEduardo Antonio Molinari NovoaAún no hay calificaciones
- Cordillera Cóndor Ecuador-Peru Jan-1997 PDFDocumento234 páginasCordillera Cóndor Ecuador-Peru Jan-1997 PDFJosé Luis Espinoza AmiAún no hay calificaciones
- The University of Dundee Botanic Garden: A Guide To The Garden and Its PurposeDocumento26 páginasThe University of Dundee Botanic Garden: A Guide To The Garden and Its PurposeChromaticghost2Aún no hay calificaciones
- Flora de Guatemala Vol.24 - Parte 3Documento454 páginasFlora de Guatemala Vol.24 - Parte 3Dana NelumboAún no hay calificaciones
- Abrams, L. 1951. Illustrated Flora of The Pacific States Vol III. GERANIACEAE To SCROPHULARIACEAE - Geraniums To FigwortsDocumento884 páginasAbrams, L. 1951. Illustrated Flora of The Pacific States Vol III. GERANIACEAE To SCROPHULARIACEAE - Geraniums To FigwortspolleiriAún no hay calificaciones
- Native Seed Mix DMDocumento83 páginasNative Seed Mix DMMeghan ManhattonAún no hay calificaciones
- Endemic Woody PlantsDocumento210 páginasEndemic Woody PlantsAnantha Krishna K SAún no hay calificaciones
- Bevis, J. F. and H. J. Jeffery. British Plants Their Biology and Ecology (1920)Documento361 páginasBevis, J. F. and H. J. Jeffery. British Plants Their Biology and Ecology (1920)shelfedgeAún no hay calificaciones
- Int Gtr277 Vascular Flora of West Central Montana Karl LackschewitzDocumento654 páginasInt Gtr277 Vascular Flora of West Central Montana Karl LackschewitzkerizlAún no hay calificaciones
- Natural History of Ceylon: The ElephantDocumento515 páginasNatural History of Ceylon: The ElephantGutenberg.orgAún no hay calificaciones
- Forest Restoration in LandscapesDocumento440 páginasForest Restoration in Landscapesleguim6Aún no hay calificaciones
- Forest Planting 1893Documento266 páginasForest Planting 1893OceanAún no hay calificaciones
- Mora Urpi Et Al 1997Documento83 páginasMora Urpi Et Al 1997Patrícia NazárioAún no hay calificaciones
- Sankaran, Mahesh - Scogings, Peter F - Savanna Woody Plants and Large Herbivores (2020, John Wiley & Sons LTD)Documento781 páginasSankaran, Mahesh - Scogings, Peter F - Savanna Woody Plants and Large Herbivores (2020, John Wiley & Sons LTD)Ariadne Cristina De AntonioAún no hay calificaciones
- Plant Invaders of Mid-Atlantic Natural AreasDocumento172 páginasPlant Invaders of Mid-Atlantic Natural AreasMaryland Native Plant ResourcesAún no hay calificaciones
- Portland Plant ListDocumento202 páginasPortland Plant ListdiccfishAún no hay calificaciones
- Identification of Pollen Allelochemical in Hieracium X Dutillyanum Lepage and Its Ecological Impacts On Conyza CanadensisDocumento1 páginaIdentification of Pollen Allelochemical in Hieracium X Dutillyanum Lepage and Its Ecological Impacts On Conyza Canadensisbrkica2011Aún no hay calificaciones
- CarobDocumento92 páginasCarobJaga Deepan100% (1)
- North Pacific Temperate Rainforests: Ecology and ConservationDocumento31 páginasNorth Pacific Temperate Rainforests: Ecology and ConservationUniversity of Washington PressAún no hay calificaciones
- An Island Biogeographical Approach To The Analysis of Butterfly Community Patterns in Newly Designed ParksDocumento2 páginasAn Island Biogeographical Approach To The Analysis of Butterfly Community Patterns in Newly Designed ParksElpe Bibas0% (1)
- Direct Root Tip Conversion of Catasetum Into Protocorn-Like BodiesDocumento6 páginasDirect Root Tip Conversion of Catasetum Into Protocorn-Like BodiesAdarilda BenelliAún no hay calificaciones
- Directory of Protected Areas in East Asia (IUCN 2002)Documento109 páginasDirectory of Protected Areas in East Asia (IUCN 2002)Victor CiocalteaAún no hay calificaciones
- The Flora of British IndiaDocumento725 páginasThe Flora of British IndiaPradeepa SerasingheAún no hay calificaciones
- Ecological Surveys of Forest Service Research Natural Areas in CaliforniaDocumento183 páginasEcological Surveys of Forest Service Research Natural Areas in CaliforniaPACIFIC SOUTHWEST RESEARCH STATION REPORTAún no hay calificaciones
- Tropical Woods 1926-5-8Documento85 páginasTropical Woods 1926-5-8Pedro Glecio Costa LimaAún no hay calificaciones
- September 1999 North American Native Orchid JournalDocumento101 páginasSeptember 1999 North American Native Orchid JournalNorth American Native Orchid JournalAún no hay calificaciones
- The Soil Mites of the World: Vol. 1: Primitive Oribatids of the Palaearctic RegionDe EverandThe Soil Mites of the World: Vol. 1: Primitive Oribatids of the Palaearctic RegionAún no hay calificaciones
- Vegetation of Australian Riverine Landscapes: Biology, Ecology and ManagementDe EverandVegetation of Australian Riverine Landscapes: Biology, Ecology and ManagementSamantha CaponAún no hay calificaciones
- Gardeners of Eden: Rediscovering Our Importance to NatureDe EverandGardeners of Eden: Rediscovering Our Importance to NatureCalificación: 3.5 de 5 estrellas3.5/5 (2)
- Phyllostomid Bats: A Unique Mammalian RadiationDe EverandPhyllostomid Bats: A Unique Mammalian RadiationAún no hay calificaciones
- Bat Roosts in Rock: A Guide to Identification and Assessment for Climbers, Cavers & Ecology ProfessionalsDe EverandBat Roosts in Rock: A Guide to Identification and Assessment for Climbers, Cavers & Ecology ProfessionalsAún no hay calificaciones
- Atlas of Pyrenulaceae and Trypetheliaceae Vol 4: Lichenized AscomycotaDe EverandAtlas of Pyrenulaceae and Trypetheliaceae Vol 4: Lichenized AscomycotaAún no hay calificaciones
- 1 Kahoot Workshop For Teachers Lesson PlanDocumento3 páginas1 Kahoot Workshop For Teachers Lesson Planapi-355425590Aún no hay calificaciones
- CHAPTER 1: Introduction To Technology For Teaching and Learning Lesson 3: Roles of Technology For Teaching and LearningDocumento2 páginasCHAPTER 1: Introduction To Technology For Teaching and Learning Lesson 3: Roles of Technology For Teaching and LearningErika Mae TupagAún no hay calificaciones
- Beeg Cover Letter and ResumeDocumento3 páginasBeeg Cover Letter and Resumeapi-252522445Aún no hay calificaciones
- Effective Teacher EssayDocumento4 páginasEffective Teacher Essayapi-644993636Aún no hay calificaciones
- Prof ReadingDocumento2 páginasProf ReadingCynthia Unay100% (1)
- AP 8 Reviewer 2Documento2 páginasAP 8 Reviewer 2lucdahAún no hay calificaciones
- PROF Ed 105: Assessment in Learning 1Documento15 páginasPROF Ed 105: Assessment in Learning 1scarsofthemooonnnnAún no hay calificaciones
- The Effectiveness of Problem-Based Learning On Students' Problem Solving Ability in Vector Analysis CourseDocumento7 páginasThe Effectiveness of Problem-Based Learning On Students' Problem Solving Ability in Vector Analysis CourseVIRA ZANUBA KHOFSYAH Tarbiyah dan KeguruanAún no hay calificaciones
- PHD Thesis - Irfan AhmedDocumento215 páginasPHD Thesis - Irfan AhmedDr Irfan Ahmed RindAún no hay calificaciones
- Reflective Report On Micro TeachingDocumento4 páginasReflective Report On Micro TeachingEzzah Syahirah100% (2)
- Treasure Hunt PBL UnitDocumento21 páginasTreasure Hunt PBL Unitapi-272198179Aún no hay calificaciones
- Stirrers and Settlers For The Primary Classroom - TeachingEnglish - British Council - BBCDocumento5 páginasStirrers and Settlers For The Primary Classroom - TeachingEnglish - British Council - BBCTomika RodriguezAún no hay calificaciones
- Lingvist Blended Learning GuideDocumento14 páginasLingvist Blended Learning GuideCarlos NicolásAún no hay calificaciones
- Rpms Teacher 1-3Documento10 páginasRpms Teacher 1-3Geoffrey MilesAún no hay calificaciones
- Teacher Communication Letter 1 FinalDocumento3 páginasTeacher Communication Letter 1 Finalapi-402035634Aún no hay calificaciones
- TSCI Paper Educ516Documento12 páginasTSCI Paper Educ516james100% (1)
- Guide To Writing Instructional Objectives in A Standards-Based SystemDocumento13 páginasGuide To Writing Instructional Objectives in A Standards-Based SystemMaria Cristina ImportanteAún no hay calificaciones
- February 18 Issue 2011Documento36 páginasFebruary 18 Issue 2011BlakeBeatAún no hay calificaciones
- Tutorial Letter 102/0/2023: Teaching Technology in The Senior PhaseDocumento14 páginasTutorial Letter 102/0/2023: Teaching Technology in The Senior Phasefikile shangeAún no hay calificaciones
- Peter Chalmers ResumeDocumento1 páginaPeter Chalmers Resumeapi-487469597Aún no hay calificaciones
- Danielson Aligned Lesson Plan Template For Formal ObservationsDocumento10 páginasDanielson Aligned Lesson Plan Template For Formal Observationsapi-272846979Aún no hay calificaciones
- Talk To Me App EvalDocumento3 páginasTalk To Me App Evalapi-289801836Aún no hay calificaciones
- TC30/90 Application Form To Renew Your Full Practising Certificate ForDocumento6 páginasTC30/90 Application Form To Renew Your Full Practising Certificate ForsajdhAún no hay calificaciones
- DGKDocumento48 páginasDGKShahbaz Hassan WastiAún no hay calificaciones
- Date / Day Time / Class Focus Theme Topic Content Standard Learning Standard Learning Objectives /S/(S) /æ/ (A) /T/ (T) /P/ (P)Documento7 páginasDate / Day Time / Class Focus Theme Topic Content Standard Learning Standard Learning Objectives /S/(S) /æ/ (A) /T/ (T) /P/ (P)Nieyfa RisyaAún no hay calificaciones
- 2 How World Religions BeganDocumento2 páginas2 How World Religions BeganGrachelle Janette Hidalgo ValienteAún no hay calificaciones
- 503 - Comprehensive ExaminationDocumento4 páginas503 - Comprehensive ExaminationJomaj Dela CruzAún no hay calificaciones
- Culturally Responsive TeachingDocumento9 páginasCulturally Responsive Teachingapi-300854245Aún no hay calificaciones
- Final Paper Chpater 1 3Documento49 páginasFinal Paper Chpater 1 3Carl Vincent SorianoAún no hay calificaciones
- EL 100 Msjbfinal OBE Intro. To LinguisticsDocumento9 páginasEL 100 Msjbfinal OBE Intro. To LinguisticsJovilleAún no hay calificaciones